 “Aun tenemos en nuestras cabezas estructuras cerebrales muy parecidas a las del caballo y el cocodrilo“, dice el neurofisiólogo Paul MacLean, del National Institute of Mental Health (NIMH). No sé si habréis oído hablar del complejo “R” o cerebro reptiliano.
“Aun tenemos en nuestras cabezas estructuras cerebrales muy parecidas a las del caballo y el cocodrilo“, dice el neurofisiólogo Paul MacLean, del National Institute of Mental Health (NIMH). No sé si habréis oído hablar del complejo “R” o cerebro reptiliano.
De acuerdo a la teoría de MacLean, el cerebro humano resulta de la superposición e integración de las funciones de tres cerebros distintos, con diferentes características estructurales, neurofisiológicas y de comportamiento.
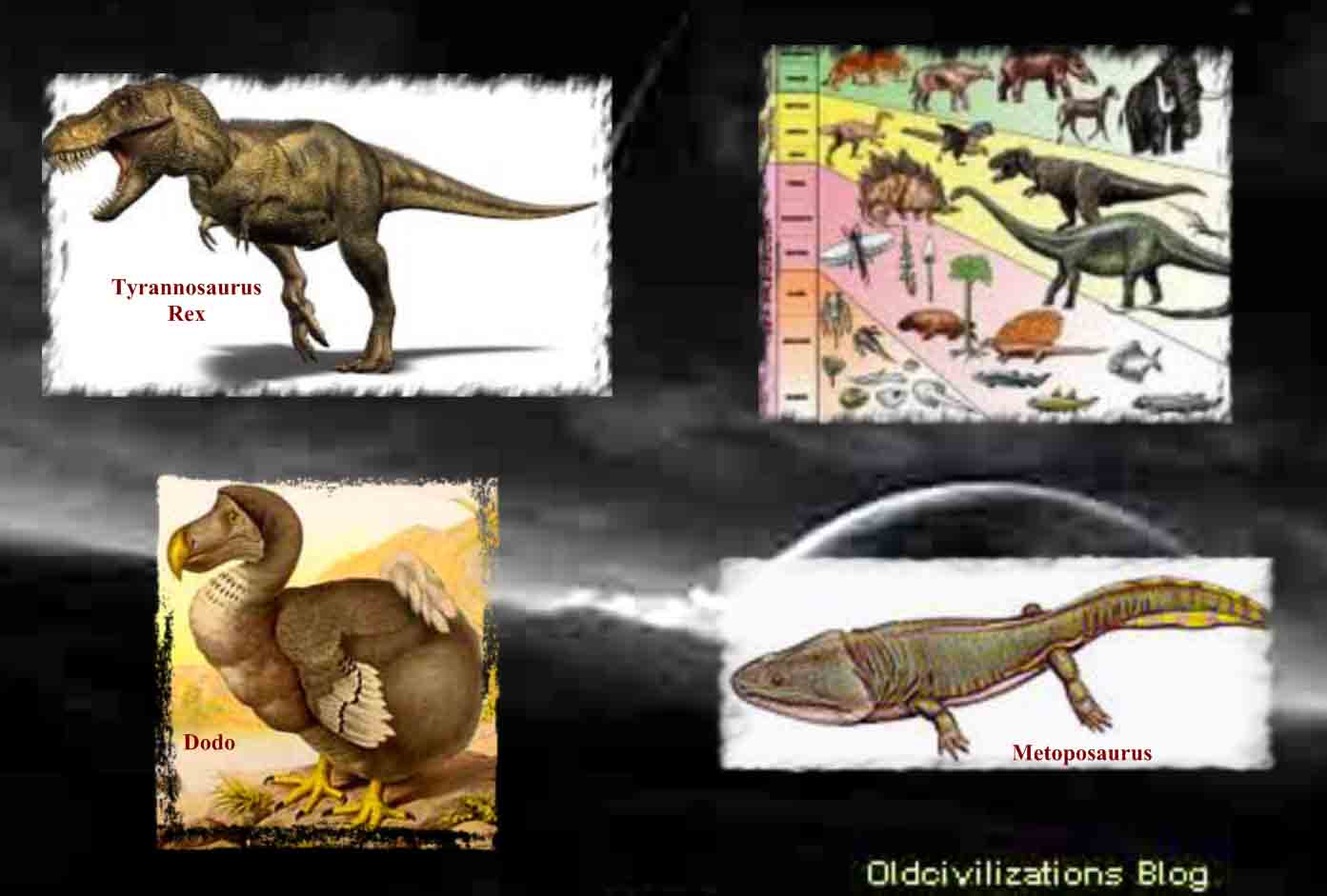
A lo largo de su evolución, el cerebro humano adquirió tres componentes que fueron surgiendo y superponiéndose. El primero es el cerebro primitivo (arquipálio), constituido por la estructuras del tronco cerebral: Bulbo, cerebelo, ponte y mesencéfalo, con el más antiguo núcleo en la base, el globo pálido y bulbos olfatorios. Se dice que corresponde al cerebro reptiliano, también llamado complejo-R por el neurofisiologo Paul MacLean. El segundo es el cerebro intermedio (paleopálio), formado por las estructuras del sistema límbico. Se dice que corresponde al cerebro de los mamíferos inferiores. El tercero, en la capa superior, es el cerebro superior o racional (neopálio), que comprende la mayor parte de los dos hemisferios cerebrales (formado por el neocórtex) y algunos grupos neuronales subcorticales. Este último solo es compartido por los mamíferos superiores, incluyendo a los primates y el hombre. También son muy comunes los mitos sobre la “Serpientes de la sabiduría” que iluminaban a la humanidad. En el libro de Génesis, Dios castiga a la serpiente por haberle dado a Eva la fruta prohibida declarando; “sobre tu vientre andarás y polvo comerás todos los días de tu vida” (Génesis 3). De esta manera la Biblia daba a entender que la serpiente, antiguamente, tenia brazos y piernas, y que, al ser maldecida, le fue quitado este privilegio. También en las viejas catedrales europeas se ven adornos de gárgolas, que son de tipo reptiloide. Asimismo, también hay muchas referencias al cruce entre “dioses” de origen reptiliano, como los anunnaki, y humanos, así como a manipulaciones genéticas con humanos, lo que aún fortalecería más la idea del posible origen de esta parte reptiliana del cerebro. Los paleontólogos, desde principios del siglo XIX, han tratado de resolver el enigma de los dinosaurios, el conjunto de especies que han dominado nuestro planeta durante un período de 165 millones de años. Revolucionando las antiguas teorías sobre estos animales, descubrimientos recientes han probado que no todos ellos eran lentos, de sangre fría, sino que algunas especies eran de sangre caliente y de movimientos rápidos, y que su desaparición no se debió a su incapacidad de adaptación, sino a grandes cataclismos que asolaron la Tierra. También se ha confirmado que diversos tipos de dinosaurio eran bípedos, como los primates y las aves. Tal vez algunas especies reptiles alcanzaron, hace más de 65 millones de años, un gran desarrollo tecnológico, que les permitiese huir de la Tierra ante un inminente cataclismo. ¿Serán descendientes de estos reptiles terrestres los “dioses” anunnaki que intervinieron en nuestra evolución? Conozcamos más sobre estos posibles ancestros: los dinosaurios.

Desde que a principios del siglo XIX fueron identificados por primera vez sus restos, los dinosaurios no sólo han apasionado a los científicos sino que han logrado fascinar al gran público. Pero, realmente, ¿qué sabemos de estos animales que, tras dominar durante 165 millones de años la vida de nuestro planeta, desaparecieron hace ahora unos 65 millones de años, al parecer de «muerte súbita»? Para que pudiera nacer la Paleontología primero fue necesario que se admitiera que la Tierra era lo suficientemente antigua como para que, durante su existencia, hubieran podido aparecer y desaparecer especies de las que el hombre nunca tuvo noticia directa. Las reconstrucciones realizadas a partir de los primeros fósiles de dinosaurios presentaban a unos enormes y pesados lagartos, provistos de unas extremidades posteriores desproporcionadas que les daban, vistos desde atrás, la apariencia de gigantescas ranas. Pero la multiplicación de los hallazgos y los avances de la investigación fueron permitiendo a los científicos aproximarse a una realidad plural. Como es lógico, a lo largo de 165 millones de años de existencia, los dinosaurios evolucionaron, se diversificaron y se adaptaron a condiciones ambientales cambiantes. Existieron especies enanas y otras gigantescas. Animales bípedos y cuadrúpedos. Herbívoros, insectívoros y carnívoros. Algunas familias estaban provistas de plumas, aunque no volaban, y otras volaban, pero no poseían plumas. Ágiles y veloces a pesar de su soberbia envergadura, algunas especies debían tener necesariamente sangre caliente. El gran enigma de los dinosaurios es su final. Hay algunas hipótesis que mantienen que no hubo una extinción total, sino una evolución hacia formas mejor adaptadas, como las aves. Después de la hegemonía de las teorías gradualistas para explicar los cambios geológicos, climáticos y biológicos producidos en la Tierra, descubrimientos de origen diverso volvieron a poner de actualidad, desde mediados de la década de los sesenta, las teorías catastrofistas. Una familia de científicos, los Álvarez, se dedicaron a la búsqueda de pruebas que avalaran su hipótesis de que un asteroide chocó con la Tierra, hace unos 65 millones de años, provocando, entre otras catástrofes, la extinción de los dinosaurios.
Los continentes habían comenzado a separarse siguiendo un camino de disgregación. El más reciente de los continentes, Pangea, se dividió en dos masas terrestres, Laurasia, en el norte, y Gondwana, en el sur, así como en otros fragmentos con las formas aproximadas de los continentes actuales. Norteamérica se desplazaba hacia el oeste y hacia el norte. Un amplio golfo acuático, el modesto principio del actual océano Atlántico, separaba a América de las tierras del este que estaba dejando atrás. Los grandes trozos de tierra que habrían de convertirse en Europa y Asia se dirigían hacia el este. África se separaba de América del Sur, la Antártida de África y Australia de la Antártida. Pero no todos los lazos quedaron cortados. Durante mucho tiempo las criaturas aún podrían emigrar por istmos y brazos de tierra entre Europa y América, o entre América y Asia. En términos generales el mundo era más cálido en aquel tiempo. Los polos estaban libres de hielo. El coral crecía en las templadas aguas de Europa, en aquel entonces más un archipiélago que un continente. En la cuenca subtropical de América del Norte los ríos fluían cruzando amplias llanuras de aluvión. Un mar cálido y poco profundo penetraba por el sudeste, donde en la actualidad están los Estados de la Costa del Golfo y durante millones de años el mar fue dividiendo el continente, principalmente en la región que hoy ocupan las Grandes Praderas americanas. A lo largo de las orillas del mar y de los ríos, por todo el país florecía la vida. Todo fue terroso y verde hasta hace sólo unos cien millones de años, cuando las plantas con flores, las angioespermas, iniciaron su evolución. Introdujeron el color de la primavera entre las cicadas y las coniferas de hoja ancha, entre las libélulas y los reptiles voladores, entre las tortugas, los lagartos y los cocodrilos, entre los pequeños mamíferos, parecidos a los ratones, que se ocultaban en el follaje y entre quienes entonces eran los señores de la vida: los dinosaurios. Éstos eran los mayores animales de aquella época conocida como la Era de los Reptiles. Los dinosaurios surgieron, a partir de sus primitivos antepasados reptiles, en algún momento del período triásico del tiempo geológico, que se extiende desde hace unos doscientos cuarenta y ocho millones a doscientos trece millones de años. Fueron los animales dominantes en el jurásico, durante los siguientes setenta millones de años, y continuaron vigorosamente hasta cerca del cretáceo, hace unos sesenta y cinco millones de años aproximadamente. Su extinción en ese período, conjuntamente con la muerte de un importante número de otras criaturas y plantas, define el fin del cretáceo para los geólogos que descubrieron un cambio abrupto en los registros de los fósiles que señalaba uno de los límites cronológicos más marcados desde la perspectiva biológica.
Cada uno de esos períodos está caracterizado por capas de rocas muy distintas y definidas que contienen una acumulación de fósiles únicos. Así es cómo los geólogos llegan a determinar el tiempo relativo en la historia de la tierra y cómo los paleontólogos pueden decir cuándo aparecieron algunas especies de dinosaurios, u otras formas de vida, y cuándo desaparecieron. El triásico, el jurásico y el cretáceo son subdivisiones de ese extenso marco cronológico conocido como el mesozoico, la era que seguramente marca el “ecuador” en la historia de la Tierra. El mesozoico se encuentra entre el paleozoico, que comenzó hace quinientos noventa millones de años, con la presencia de fósiles en abundancia, y el cenozoico, la era de los nuevos animales, que cubre los últimos sesenta y cinco millones de años. La Era Mesozoica, Mesozoico, o Era Secundaria, conocida zoológicamente como la era de los dinosaurios o botánicamente como la era de las cícadas, es una división de la escala temporal geológica que se inició hace 251 millones de años y finalizó hace 65 millones de años. Se denomina Mesozoico porque se encuentra entre las otras dos eras del eón Fanerozoico, la era Paleozoica y la era Cenozoica. Durante estos 186 millones de años no se produjeron grandes movimientos orogénicos. Los continentes gradualmente van adquiriendo su configuración actual. El clima fue excepcionalmente cálido durante todo el período, desempeñando un papel importante en la evolución y la diversificación de nuevas especies animales. Los invertebrados característicos de este período fueron los amonites, de caparazón con forma de caracol, y los belemnites, más pequeños y con el caparazón alargado y puntiagudo, entre otros equinodermos, braquiópodos y cefalópodos. Se desarrollaron ampliamente los vertebrados, sobre todo los reptiles, por lo que a la Era Secundaria se le llama también la Era de los reptiles o Era de los Dinosaurios. En esta era aparecen también los mamíferos, las aves y las angiospermas o plantas con flores. El límite inferior del Mesozoico viene fijado por la extinción masiva del Pérmico-Triásico, durante la cual aproximadamente del 90% a 96% de las especies marinas y el 70% de los vertebrados terrestres se extinguieron. También es conocido por la “Great Dying“, ya que es considerada la extinción masiva más grande de la historia. El límite superior se fija en la extinción masiva del Cretácico-Terciario, que pudo haber sido causada por el impacto que creó el Cráter de Chicxulub en la Península de Yucatán. Aproximadamente el 50% de todos los géneros se extinguieron, incluidos todos los dinosaurios no avianos, es decir, todos aquellos que no compartían las características que definen a un ave. En comparación con la vigorosa convergencia de placas formadora de montañas de finales de la Era Paleozoica, las deformaciones tectónicas del Mesozoico fueron relativamente leves. Sin embargo, la era destaca por la gran fragmentación del supercontinente Pangea. Pangea gradualmente se dividió en un continente norte, Laurasia y un continente sur, Gondwana con apertura del océano Atlántico. Esto creó el margen continental que caracteriza a la mayor parte de la costa atlántica actual.

A finales de la era, los continentes se habían fragmentado a casi su forma actual. Laurasia se convirtió en América del Norte y Eurasia, mientras que Gondwana se dividió en América del Sur, África, Australia, Antártida y el subcontinente indio, que colisionará con la placa asiática durante el Cenozoico. La formación de la cordillera de los Andes comenzó en el Jurásico, pero fue durante el Cretácico cuando tomó su forma actual. Se debió al movimiento de subducción de la placa de Nazca por debajo de la placa Sudamericana. A finales del Cretácico comenzó la orogenia Laramide, que continuaría durante la primera mitad del Cenozoico y que formó las montañas Rocosas. El Mesozoico concluyó con el impacto de un asteroide de unos 10 kilómetros de diámetro, que ocasionó la extinción masiva del Cretácico-Terciario. El Triásico fue en general seco, una tendencia que comienza a finales del Carbonífero, y muy estacional, especialmente en el interior de Pangea. El bajo nivel del mar también puede haberse exacerbado por las temperaturas extremas. Con su alto calor específico, el agua actúa como un estabilizador de temperatura y un reservorio de calor, por lo que las tierras próximas a las grandes masas de agua, especialmente de los océanos, experimentan una menor variación de temperatura. Dado que gran parte de las tierras que constituían Pangea estaban lejos de los océanos, las temperaturas fluctuaban mucho, y en el interior de Pangea probablemente incluía extensas zonas desérticas. Se dispone de abundantes camas rojas y evaporitas, tales como sales, que apoyan estas conclusiones. El nivel del mar comenzó a subir durante el Jurásico, lo que probablemente fue causado por una expansión del lecho marino. La formación de nueva corteza bajo la superficie desplazó las aguas oceánicas hasta 200 m más que actualmente, lo que inundó las zonas costeras. Por otra parte, Pangea comenzó a romperse en fragmentos más pequeñas, con lo que el aumento de superficie en contacto con el océano formó el Mar de Tetis. La temperatura continuó aumentando y comenzó a estabilizarse. La humedad también aumentó con la proximidad del agua y los desiertos se retiraron. El clima del Cretácico es menos conocido y más ampliamente discutido. Se cree que los niveles más elevados de dióxido de carbono en la atmósfera causaron un gradiente de temperatura de norte a sur casi plano: las temperaturas son más o menos las mismas en todo el planeta. Las temperaturas medias son también más elevadas que en la actualidad, alrededor de 10 °C más. De hecho, a mediados del Cretácico, las aguas ecuatoriales del océano (quizás tan cálidas como 20 °C en las profundidades del océano) pueden haber sido demasiado altas para la vida marina, y las zonas terrestres cerca del ecuador pueden haber sido desiertos, a pesar de su proximidad al agua. La circulación de oxígeno a las profundidades del océano también puede haberse interrumpido.
Por esta razón, los grandes volúmenes de materia orgánica acumulada que no podían descomponerse y fueron depositados como “pizarras“. No todos los datos apoyan estas hipótesis, sin embargo. Incluso con el calentamiento, las fluctuaciones de temperatura deberían haber sido suficiente para la formación de casquetes polares y glaciares, pero no hay pruebas de ninguno de ellos. Los modelos cuantitativos también han sido incapaces de recrear la planitud del gradiente de temperatura del Cretácico. La extinción de casi todas las especies animales al final del Pérmico permitió la radiación de numerosas formas de vida nuevas. En particular, la extinción de los grandes herbívoros y los carnívoros Dinocephalia dejaron vacíos estos nichos ecológicos. Algunos fueron ocupados por los cinodontos y dicinodontos sobrevivientes, los últimos de los cuales posteriormente se extinguieron. Sin embargo, la fauna del Mesozoico estuvo dominada por los grandes arcosaurios que aparecieron unos pocos millones de años después de la extinción masiva del Pérmico-Triásico: dinosaurios, pterosauros y los reptiles acuáticos como ictiosaurios, plesiosauro y mosasauros. Los cambios climáticos de finales del Jurásico y Cretácico proporcionaron más condiciones favorables para la radiación adaptativa. En el Jurásico se produjo la mayor diversidad de los arcosaurios, y cuando aparecieron las primeras aves y mamíferos placentarios. La angiospermas se diversificaron en algún momento del Cretácico temprano, primero en los trópicos, pero el gradiente de temperatura les permitió la propagación hacia los polos a lo largo del período. Al final del Cretácico, las angiospermas dominaron la flora arbórea en muchas zonas, aunque algunas pruebas sugieren que la biomasa siguió dominado por cicas y helechos hasta después de la extinción masiva del Cretácico-Terciario. Algunos investigadores han argumentado que los insectos se diversificaron con las angiospermas, porque su anatomía, especialmente las partes de la boca, parecen particularmente bien adaptados a las plantas con flores. Sin embargo, todas las partes principales de la boca de los insectos precedieron a las angiospermas y la diversificación de insectos en realidad se redujo cuando estas surgieron, por lo que su anatomía original debe haber estado adaptada para otros fines. Cuando la temperaturas de los mares se incrementó, los animales más grandes de principios de la era mesozoica gradualmente comenzaron a desaparecer mientras que los animales más pequeños de todo los tipos, incluidos los lagartos, serpientes y quizás el antecesor de los primates, evolucionaron. La extinción masiva del Cretácico-Terciario acentuó esta tendencia.Los grandes arcosaurios se extinguieron, mientras que las aves y los mamíferos prosperaron, tal como lo hacen hoy.
Aunque los dinosaurios poblaron todas las grandes masas terrestres, con la posible excepción del continente antártico, es en la parte oeste de Norteamérica donde parecieron encontrar el hábitat más idóneo. Allí estuvieron presentes por todas partes, en una forma u otra, durante el mesozoico, desde Texas y Nuevo México hasta Alberta. Pero no todos ellos se correspondían con la creencia popular. Algunos de ellos no eran mayores que conejos o cuervos. Otros en aquella cuenca subtropical, como en muchas otras partes, llegaron a alcanzar los veinticinco o treinta metros de longitud con extravagantes colas y cuellos que abarcaban gran parte de esa longitud. Esos gigantes, Diplodocus, Brontosaurus y Brachiosaurus, son los mayores animales terrestres que jamás existieron. Algunos dinosaurios se sostenían sobre fuertes miembros traseros y, erguidos, destacaban sobre todos los demás. Tenían cuernos, garras y su cuerpo estaba protegido por una especie de armadura, como si estuvieran preparados para un combate a muerte. El Tyrannosaurus rex tenía dos patas y una cabeza excepcionalmente grande para un dinosaurio. Sus afilados dientes eran muy grandes, lo que debía de hacer que su presencia resultara espantosa a finales del cretáceo. Algunos otros tenían picos parecidos a los de los patos. Muchos tenían las patas semejantes a las aves actuales. Algunos corrían como los avestruces, mientras que otros eran perezosos. Unos eran herbívoros, pastaban, aunque no había nada de pastoral en un Triceratop cuando atacaba, cargando como un rinoceronte. Otros eran carnívoros y cazaban en manadas. Unos trescientos cuarenta géneros de dinosaurios, grandes y pequeños, fieros y dóciles, bípedos y cuadrúpedos, herbívoros y carnívoros, han sido identificados partiendo de sus restos fosilizados no sólo en Norteamérica sino en todo el mundo. Ningún género sobrevivió durante toda la Era de los Reptiles, pero los dinosaurios, de un tipo o de otro, vivieron durante ciento sesenta millones de años y los últimos de ellos debieron de morir ya cerca del período cretáceo. Sus cuerpos se hundieron en los sedimentos de aquellos viejos pantanos y en las riberas de los ríos o los lagos. Sus huesos, que se convirtieron en fósiles, quedaron a la espera de ser descubiertos. Biólogos y paleoantropólogos siguen la evolución de la vida en general y del hombre en particular, una narrativa con más puntos cruciales de transición decisivos que ningún historiador clásico podría encontrar en los archivos. Los paleontólogos son, por definición, exploradores del tiempo. El nombre «paleontología» fue acuñado por Charles Lyell en 1838, como una derivación de tres palabras griegas que significan «la ciencia del ser antiguo». Los paleontólogos tratan de recrear la historia de la vida pasada a partir de los fósiles, y entre el material más importante del que se ocupan están los restos fosilizados de los dinosaurios. Los restos de los dinosaurios sólo fueron identificados por vez primera a principios del siglo XIX.
Stephen Jay Gould, paleontólogo de la Universidad de Harvard y brillante escritor de ensayos científicos, describe al paleontólogo como: «uno de esos excéntricos que hicieron de su fascinación infantil por los dinosaurios una profesión». La contemplación de un esqueleto deTyrannosaurus en un museo, una imagen pavorosa al principio, despertó en Gould una curiosidad que lo llevaría a dedicarse, de adulto, al estudio de la paleontología y de los grandes logros de la evolución. Dale A- Russell, del Museo Nacional de Ciencias Naturales del Canadá, en Otawa, explica que: «De niño podía imaginarme el mundo de los dinosaurios. A veces yo mismo era un dinosaurio. Así, cuando me hice mayor, en vez de dedicarme a algo práctico seguí ocupándome de los dinosaurios». Un siglo antes, Charles Darwin dijo: «El placer del primer día de caza no puede ser comparado con lo que significa el hallazgo de un buen grupo de huesos fósiles, que nos cuentan su historia de los tiempos pasados casi con el mismo vigor que una lengua viva». Durante varias décadas, a mediados del siglo XX, el campo de la investigación del dinosaurio pareció tan petrificado como sus huesos. Los dinosaurios se consideraban tan definitivamente extinguidos y tan estigmatizados por su irrevocable desaparición, que se creyó que bien poco podía aprenderse de ellos. Pero en la década de 1960-1970, los paleontólogos, lo mismo que los biólogos, geólogos y físicos, comenzaron a competir entre sí para introducir nuevos hallazgos e hipótesis sobre la vida y la muerte de aquellas grandes bestias. Los debates sobre esas teorías revivieron en las páginas de las publicaciones científicas y universitarias. Nuevos conocimientos sobre los dinosaurios, combinados con los adelantos en geología, bioquímica, paleomagnetismo, paleobotánica y paleografía, inspiraron nuevas ideas sobre 1a evolución y la extinción de la vida en la Tierra. Entre los años setenta y los ochenta del siglo XX, diversos descubrimientos revelaron muchos más datos sobre la variedad de los dinosaurios. En la década de los setenta se descubrieron cincuenta nuevos tipos de dinosaurios. Cuando los paleontólogos reunieron y conjuntaron los hallazgos fósiles y estudiaron su significado empezó a tomar forma una nueva imagen de los dinosaurios. Es posible que algunos dinosaurios llegaran a desarrollar metabolismos activos no reptiles. Había que dejar de pensar en ellos simplemente como si fueran lagartos arcaicos. Algunos de ellos posiblemente fueran seres sociales y padres atentos. Otros dinosaurios quizá perduren, en cierto modo, en algunas aves actuales. Además, es posible que la desaparición total del dinosaurio no se debiera tanto a sus propias limitaciones mentales y físicas como a fuerzas ambientales, incluyendo una posible catástrofe global. Los dinosaurios se desarrollaron, se adaptaron y sobrevivieron durante el increible período de ciento sesenta millones de años.
Ello seguramente provocó que los mamíferos no pudieron pasar de una modesta posición, como criaturas esencialmente nocturnas y forzadas a pasar inadvertidas, subsistiendo principalmente a base de una dieta de insectos. Por lo tanto, vale la pena saber más sobre los dinosaurios, sobre todo ahora que los exploradores han empezado a descubrir en la Tierra y en los huesos enterrados en ella un pasado de dimensiones sorprendentemente amplias. Lo maravilloso de los dinosaurios no es sólo que vivieron hace mucho tiempo, crecieron tanto y después se extinguieron, en misteriosas circunstancias, millones de años antes de que, supuestamente, los seres humanos hubieran hecho su aparición en el escenario de la vida. Al principio sólo se conocieron breves indicios sobre tiempos remotos y vida pasada. Unos pocos fragmentos de huesos y algunos dientes, incrustados en antiguos sedimentos, salieron a la luz en Inglaterra en el primer cuarto del siglo XIX. Eran huesos de criaturas muertas hacia mucho tiempo, reliquias de la vida en un pasado distante. Nadie sabia cuánto tiempo hacía que habían muerto, puesto que todavía se ignoraba hasta dónde podía extenderse aquel pasado. A juzgar por el aspecto de aquellos primeros huesos y dientes debía de tratarse de criaturas extrañas y monstruosas. Pero nadie podía saber entonces cuál era su verdadero aspecto. Fueran lo que fuesen, lo único cierto era que aquellas criaturas que pasaron a ser conocidas con el nombre de dinosaurios parecían ser distintas a cualquier forma de vida existente en la actualidad. Los fósiles que empezaban a ser encontrados en número cada vez mayor y en las formas más intrigantes, no podían ser interpretados de otro modo más que como residuos remanentes de formas de vida extinguidas. Ésta fue la conclusión a la que llegó Georges Cuvier, el más destacado de los paleontólogos de principios del siglo XIX y el primer científico que presentó pruebas detalladas y convincentes de la realidad de la extinción como un fenómeno general en la historia de la vida. Cuvier no descubrió a los dinosaurios, pero hizo mucho para crear las condiciones intelectuales que harían posible su descubrimiento.
Nadie hubiera tomado en consideración los fósiles de los dinosaurios, de no haber tenido conciencia de su extinción. La idea de que una catástrofe geológica contribuyó a las distintas oleadas de extinciones había surgido para influir no sólo en el estudio de los dinosaurios sino en la investigación de las causas y en las consecuencias de las extinciones masivas a lo largo del tiempo. En 1796 Cuvier hizo sus primeras revelaciones sobre la extinción de las especies. En 1795, los ejércitos de la Francia revolucionaria estaban luchando en los Países Bajos, y a su regreso llevaron consigo un monstruoso trofeo de guerra encontrado en la entonces pequeña ciudad de Maastricht. Se trataba de una pareja de maxilares fosilizados que medían más de un metro de longitud. Los maxilares eran los restos de alguna enorme criatura antediluviana. Todos los huesos de edad y origen indeterminado se presumía que debían de pertenecer a una época anterior al bíblico diluvio. Un cirujano militar alemán, retirado, y un coleccionista de fósiles fueron convocados para examinar los fósiles. Era el maxilar de una ballena, decidió uno. No, era un gigantesco lagarto marino, opinó otro. ¿Era posible? No, jamás se habían visto lagartos de aquel tamaño ni en el mar ni en la tierra. Como muchos otros europeos llegaron a sospechar que podía tratarse de un monstruo prehistórico, algo que posiblemente había vivido antes de Noé y, probablemente, antes de Adán, y que había dejado de existir. Cuvier examinó el maxilar, lo comparó con el de otros animales conocidos y llegó a la conclusión de que pertenecía a un lagarto que podía estar relacionado con sus parientes que aún existían en los trópicos. Sin embargo era de tamaño mucho mayor, debió de haber llevado una vida marina y se alimentaría de peces. Dado que sólo presentaba un leve parecido con las criaturas conocidas, el animal debió de haber vivido en un pasado muy remoto. Unos años más tarde, el reverendo William D. Conybeare, en Inglaterra, le dio el nombre de Mosasaurus, o «lagarto del Mosa», de acuerdo con la región donde había sido encontrado. El Mosasaurus fue el primer gigante marino del mesozoico encontrado e identificado. La correcta interpretación que dio Cuvier de aquella mandíbula, hizo que pasara a ser considerado el mejor de los expertos en grandes fósiles. Se tuvo otra prueba evidente de que no todas las especies vivas habían logrado sobrevivir hasta llegar a nuestros días, cuando en Alemania se descubrió un «animal marino desconocido» que parecía tener alas. Por medio de dibujos, Cuvier identificó al animal como un reptil volador, no marino sino terrestre y le dio el nombre de Pterodactyl, derivado de las palabras griegas que significan «ala» y «dedo». Después se empeñó en que ése era uno de los pterodáctilos que vivieron en los tiempos de los dinosaurios y que fueron los primeros verdaderos vertebrados voladores. Algunos de ellos eran tan pequeños como gorriones, mientras que otros tenían enormes alas con una envergadura de hasta diez metros. Cuvier dijo que esos animales se habían extinguido.

En 1801, Cuvier presentó su tesis de que la extinción era un hecho aparente que se daba en la naturaleza. Dijo que nada podía ser más importante que «descubrir si las especies que existieron entonces habían sido completamente destruidas o si, simplemente, se habían ido modificando de un clima a otro». Esto reflejaba el reconocimiento de Cuvier de tres explicaciones alternativas para las distintas formas de fósiles que había estado examinando: extinción, evolución y migración. Concedió que, en algunos casos, se podía aplicar la última de las explicaciones, aunque resultaba poco probable que cualquiera de aquellos grandes animales terrestres hubiera escapado a la atención de los exploradores en otros continentes. En la mente de Cuvier esto reducía a dos las posibilidades y quedaba por elegir entre extinción y evolución. Para Cuvier, todas las pruebas aportadas por los fósiles hablaban en favor de la extinción como la forma más sencilla y clara de entender lo que había sucedido a la vida prehistórica. Unos pocos científicos habían sopesado previamente la posibilidad de la extinción, sin embargo no se decidieron plenamente a aceptar la idea, por temor a entrar en contradicción con el Eclesiastés, en la Biblia: «Todo lo que Dios hace, así lo sé, tiene que ser para siempre: nada se le puede añadir ni nada puede ser excluido de ello». Es posible que pensaran que las criaturas que parecían haberse extinguido sólo hubieran desaparecido de Europa y siguieran vivas en algún lugar remoto, poco explorado, en cualquier parte del mundo. Ésa fue la teoría de la migración, expuesta también por Cuvier. Cuando Thomas Jefferson envió a Meriwether Lewís y a William Clark a su expedición al Oeste americano, se sentía seguro de que estos exploradores regresarían con informes sobre la existencia de algunas especies supervivientes. Ciertos acontecimientos, entre los que destacó el descubrimiento del celacanto en las aguas de África del Sur y de un marsupial australiano llamado Burramys parvus, animales supuestamente extinguidos que aparecieron vivos, dio nuevo impulso a la tesis contraria a la extinción. Pero cuando Cuvier, con todo el peso de su autoridad y su reputación, y con pruebas en la mano, proclamó la verdad de la extinción de las especies, casi todas las dudas desaparecieron. Cuvier no sólo hizo que los científicos aceptaran la extinción sino que además desarrolló métodos de investigación que influyeron en preparar a los científicos para que pudieran saber cómo tratar los primeros huesos de los dinosaurios. Lo que hizo Cuvier fue elevar la anatomía comparada al nivel de una ciencia que habría de servir de gran ayuda a la paleontología. En la búsqueda de fósiles había que partir de la geología para establecer su edad relativa. Pero para identificar los especímenes e interpretar su lugar en la naturaleza había que saber biología y, de manera muy especial, anatomía comparada. En ese campo de la ciencia Cuvier demostró que cada grupo importante de animales tenía su particular arquitectura corporal.
Cuvier reconoció que la especie de un animal podría ser identificada normalmente por la forma de un simple hueso o, al menos, por unos cuantos huesos. El hueso del pulgar de los pies, por ejemplo, está relacionado con el hueso del arco del pie y éste, a su vez, se relaciona con el hueso de la pantorrilla y así sucesivamente. Al establecer estas conexiones, como Cuvier supo hacer tan bien, inculcó en los paleontólogos una confianza tan grande en su capacidad analítica que se aproximaba a la certeza. Esta confianza fue reflejada por Cuvier cuando escribió: «La forma y estructura del diente regula la forma del cóndilo y de otras articulaciones del omoplato y de las garras, del mismo modo que la ecuación de una curva regula todas sus otras propiedades. Así, al comenzar nuestra investigación con el cuidadoso examen de un hueso en sí, una persona que domine de manera suficiente las leyes de la estructura orgánica, podría si así lo hiciera reconstruir la totalidad del animal al que perteneció el hueso». Cuvier pudo estar equivocado en su visión global, como puso en claro uno de los descubridores de los dinosaurios, pero eso fue la excepción. Cuvier parecía tener una facilidad casi mágica para tomar unos cuantos huesos extraños y reconstruir el esqueleto completo de una criatura desconocida, mamut o mastodonte, así como antiguas formas de rinocerontes, hipopótamos, ciervos o cocodrilos. En las manos de Cuvier la paleontología parecía más un arte que una ciencia. Pero no todo el trabajo de Cuvier consistía en las reconstrucciones en el laboratorio. A veces iba al campo, frecuentemente en compañía de Alexandre Brongniart, un geólogo minero, y juntos examinaban y reconocían una excavación que penetraba en las profundidades de una capa estratificada procedente de un pasado remoto, más antiguo cuando más diferentes eran los fósiles que allí se encontraban de los animales existentes en la actualidad. Las distintas capas o estratos mostraban cambios muy marcados en la tierra y en la vida a lo largo de los tiempos. Cuvier identificó unas ciento cincuenta especies de fósiles en la cuenca del Sena y noventa de ellos no tenían equivalentes vivos. Con esas extinciones, Cuvier razonó que una secuencia de cataclismos ampliamente extendidos condenó parte de la vida primitiva y dio nueva forma a la superficie de la tierra. La cuenca del Sena había estado, alternativamente, seca e inundada. Los estratos de los tiempos en que ocurrieron esos episodios parecían contamos una historia de vida pasada y de muerte catastrófica. Cuvier presentó su teoría sobre la historia de la Tierra en un tratado de 1812, titulado A discourse on the Revolutions of the Surface of the Globe.
De acuerdo con su teoría, el mar, o grandes inundaciones, habrían invadido repetidamente las tierras. El hielo glacial avanzó cruzando continentes que hasta entonces habían sido templados, lo cual explicaría el hallazgo de algunos animales, como Mamuts, que recientemente fueron descubiertos congelados en Rusia. Movimientos muy violentos agitaron y conmovieron la costra de la superficie terrestre. Cuvier explicó que se trató de catástrofes tan repentinas y completas, que «el hilo de las operaciones de la naturaleza fue roto por ellas». «Las catástrofes barrieron de la existencia a un sinnúmero de criaturas vivas -escribió Cuvier-. Las que vivían en tierras secas fueron ahogadas por los diluvios e inundaciones. Otras, cuyo hogar estaba en las aguas, perecieron cuando el fondo de los mares se secó repentinamente. Razas enteras se extinguieron, dejando sólo simples rastros de su existencia que ahora son difíciles de reconocer incluso por los naturalistas». Con su descubrimiento de la extinción, y de la existencia de un pasado que, por lo general, es muy distinto del presente, Cuvier preparó las mentes del siglo XIX para el pensamiento de que la historia de la Tierra se extendía en el pasado hasta los tiempos anteriores a Adán y para esperar que la Tierra descubriera, bajo sus capas, más huesos de criaturas tan fabulosas como el Mosasaurus. Varios descubrimientos recientes habían llamado la atención de los científicos y del público. Todos hablaban del increíble reptil que Mary Anning había encontrado en los acantilados de Lyme Regis, en el sur de Inglaterra. Ella y su hermano descubrieron algunos huesos que sobresalían de las rocas en los acantilados y después de romper las rocas con martillos y cinceles hallaron la marca de un esqueleto de diez metros de longitud. La criatura con sus cuatro patas con garras y sus grandes mandíbulas llenas de fuertes dientes muy afilados, tenía un aspecto combinado de reptil y pez. Cuando se conoció la noticia de su hallazgo, los sabios de Londres se dirigieron en peregrinaje a la modesta vivienda de Anning. Ella había encontrado el primer esqueleto razonablemente completo de un reptil marítimo que más tarde pasaría a ser conocido con el nombre de Ichthyosaurus, término derivado de las palabras griegas que significan «pez» y «lagarto». Unos años más tarde la propia Mary Anning descubriría los restos de otro reptil marino, el Plesiosaurus. Gideon Algemon Mantell comenzó la búsqueda de fósiles y dio con los huesos y dientes de la que resultó ser la más famosa víctima de las extinciones deducidas por Cuvier. En un día de primavera de 1822, Mantell fue en un coche de caballos hasta el bosque de las afueras de Lewes, en Sussex, donde el río Ouse transcurre hacia el sur, en dirección al canal de la Mancha. Allí, Mantell se detuvo en una casa para visitar a un paciente. Mientras estuvo allí, su esposa, Mary Ann, lo esperaba paseando por la carretera.

La esposa de Mantell había llegado a participar en el interés de su marido por los fósiles. Habían colaborado en un extenso libro, The Fossils of the South Downs, que debía ser publicado en ese mismo año, en el cual reproducía su colección de conchas marinas fósiles, la mayor parte de ellas muy antiguas. Consecuentemente no había nada más natural para Mary Ann Mantell que fijarse en un montón de piedras dejado allí por los peones camineros que habían reparado la carretera. Sus ojos se fijaron principalmente en un trozo de piedra arcillosa. Encerrado en ella había algo que a primera vista podría tomarse por un gran diente. Se lo enseñó a su marido tan pronto éste salió de la casa. «Has encontrado los restos de un animal hasta ahora desconocido por la ciencia», se supone que Mantell le dijo a su mujer. El diente, según diría después, era «totalmente distinto a cualquier otro que hubiera visto con anterioridad». Mantell visitó varias canteras y pidió a los picapedreros que le avisaran si encontraban algún otro diente o cualquier otra cosa que fuera más allá de las conchas normales. Aparecieron varios dientes más, así como algunos extraños huesos en una cantera de Tilgate Forest, cerca de Cuckfield, en Sussex. Resultaron ser los huesos y los dientes del primer dinosaurio reconocido como tal. Pese a que la importancia de ese descubrimiento pasó inadvertida al principio, mientras Mantell examinaba y admiraba cada vez más esos dientes, se sentía intrigado. Notó que la corona de uno de los dientes estaba desgastada y formaba una superficie oblicua y suave. Estaba claro que se trataba de un animal de gran tamaño que se alimentaba de plantas. Aquel diente no podía haber masticado carne. El diente le recordaba a Mantell «parte de un incisivo, desgastado por el uso, de un gran paquidermo». Pero él sabía, gracias a los descubrimientos de Cuvier de algunas especies de elefantes extintas, que los restos de esos mamíferos prehistóricos sólo se encontraban en los estratos superiores pertenecientes a épocas relativamente recientes y no en los estratos más antiguos, más bajos, de los cuales creía que provenía su diente. Consecuentemente los pensamientos de Mantell se volvieron de nuevo a los reptiles antiguos. Dado que la edad aparente del diente fósil de Mantell parecía excluir cualquier conexión con los mamíferos, Mantell creía que debía de tratarse de reptiles. Pero una característica de los reptiles modernos no se acomodaba con esa línea de pensamiento. «Como ninguno de los reptiles existentes son capaces de masticar sus alimentos -dijo Mantell-, no podía arriesgarme a atribuir el diente en cuestión a un saurio». Mantell tomó aquel diente y otros ejemplares y se los llevó consigo a una reunión de la Sociedad Geológica de Londres. Pero sólo el químico William Wollaston apoyaba a Mantell en su opinión de que había descubierto dientes de un «desconocido reptil herbívoro».
Mantell se enteró de que Charles Lyell, un joven abogado y geólogo, visitaría París y le pidió que se llevara consigo el diente fósil para enseñárselo a Cuvier. Lyell visitó a Cuvier en su despacho-estudio del Museo de Historia Natural. Cuvier examinó el diente y, sin la menor vacilación, dijo que se trataba simplemente de un incisivo superior de rinoceronte. Mantell insistió, pese a las opiniones de la Sociedad Geológica y de Cuvier. William Buckland, profesor de geología en la Universidad de Oxford, le escribió a Mantell pidiéndole que no publicara nada en lo que se afirmara que los fósiles provenían de capas más profundas que las superficies diluvianas -es decir, de los sedimentos superiores-. No había pruebas, dijo Buckland, de que los fósiles fuesen lo suficientemente antiguos como para pertenecer a un reptil gigante extinguido. Pese a todo, Mantell siguió insistiendo. Al parecer Mantell llevó los fósiles al Hunterian Museum de Londres. Allí se pasó muchas horas volcado sobre la colección de dientes y huesos de reptiles con la esperanza de encontrar algo comparable a los que obraban en sus manos. Su búsqueda resultó infructuosa. Sin embargo, por casualidad, aquel día se encontraba en el museo un joven llamado Samuel Stutchbury, que había estado realizando investigaciones sobre las iguanas. Cuando Mantell le enseñó sus dientes fósiles, Stutchbury observó que había una gran semejanza entre ellos y los dientes de las iguanas de América Central. Cuando Mantell contempló los especímenes de Stutchbury, también observó la semejanza. Mantell sintió renacer su confianza en que los dientes y huesos en su poder eran los de un reptil gigantesco que se alimentaba con plantas. Y si una iguana tenía dientes de aquel tamaño, especuló, el animal debió de haber sido enorme, unos dieciocho metros de longitud. Mantell le dio a aquel reptil fósil el nombre de Iguanodon, lo que significa «diente de iguana». En 1825 ofreció un completo informe de su descubrimiento a la Royal Society en Londres. Algunas de las presunciones de Mantell sobre el Iguanodon resultaron equivocadas. Pensó que el animal había caminado sobre cuatro patas a la manera de una iguana a escala mayor. Confundió su protuberancia puntiaguda encima del hocico tomándola por un cuerno y así lo hizo ver en su dibujo, lo que dio al animal cierta apariencia de rinoceronte. Pero en lo que Mantell no se equivocó fue en la importancia de su descubrimiento. Mucho tiempo atrás, antes de que los mamíferos florecieran en la Tierra, declaró, habían vivido en ésta reptiles más gigantescos que cualquiera otros que, por lo que hasta ahora sabemos, hayan existido en nuestro planeta.
Cuando Cuvier se enteró de los hallazgos de Mantell antes de su publicación, reconoció sus anteriores errores y expuso algunas ideas propias al respecto. Dado que todos los mayores animales terrestres contemporáneos eran herbívoros, era razonable pensar que los mayores entre los antiguos reptiles se «habían alimentado de vegetales». Consecuentemente no debía sorprender a nadie que el Iguanodon, siendo tan grande, tuviera dientes de animal herbívoro. Como Cuvier adelantó, y revelaron hallazgos subsiguientes, muchos de los dinosaurios resultaron ser herbívoros, contrariamente a lo que ocurre con los reptiles de nuestros días. Mientras Mantell estaba sufriendo al pensar qué debía hacer con los fósiles del Iguanodon, entre 1820 y 1825, un cazador de fósiles de los más eminentes, William Buckland, examinó en el museo de Oxford una interesante colección de huesos que habían sido encontrados en una cantera de pizarra. La mayoría de las tejas de pizarra de la región provenían de la aldea de Stonesfield, al norte de Oxford, y en los últimos años, a medida que la cantera se iba haciendo más profunda, los trabajadores fueron encontrando un número cada vez mayor de huesos fósiles que entregaron al museo. Algunos eran reliquias procedentes de pequeños mamíferos. Otros parecían ser mucho más antiguos y mayores. Entre esos huesos se encontraba un maxilar inferior con grandes dientes puntiagudos, como de sierra, algunas vértebras, un trozo de omoplato y varios fragmentos de un miembro trasero. Buckland decidió que era muy posible que los huesos procedieran de reptiles. También él estaba influenciado por el Mosasaurus de Cuvier. Por alguna razón, Buckland se tomó mucho tiempo para informar sobre sus fósiles. Pero consultó a amigos y colegas a los que hizo observar su material para que le dieran su opinión. Una de esas personas se sintió tan impresionada por los grandes dientes en forma de hoja que publicó en 1822 la declaración de que el diente de Stonesfield pertenecía a un reptil de gran tamaño al que dio el nombre de Megalosaurus. El autor fue James Parkinson, un médico que es recordado por la descripción de la enfermedad que lleva su nombre. Buckland esperó dos años para ofrecer la primera descripción completa del Megalosaurus que apareció en 1824 en una publicación científica bajo el título de Transactions of the Geological Society of London. Con esto se adelantó a Mantell en un año, aun cuando él mismo, a su vez, estuvo precedido por Parkinson. Pero Buckland fue el primero en publicar un informe científico formal sobre los restos de un dinosaurio. El artículo publicado por Mantell sobre el Iguanodon se publicó un año después, en 1825, pero había expuesto y discutido sus hallazgos ante la Geological Society meses antes de la publicación del informe de Buckland y describió ya los dientes en su libro de 1822 sobre fósiles.

La influencia de Cuvier también se hizo evidente en la obra de Buckland sobre el Megalosaurus.Parece ser que Cuvier insistió a Buckland para que siguiera adelante y publicara su artículo titulado: «Notice on the Megalosaurus or Great Fossil Lizard of Stonesfield». Al igual que Cuvier en su análisis sobre el Mosasaurus, Buckland clasificó confiadamente los fósiles en el orden de los «saurios o lagártidos». Más aún, al reconocer su deuda académica con Cuvier, escribió: «De estas dimensiones, comparadas con las medidas de la familia de los lagartos, resulta que Cuvier ha asignado al individuo al que pertenece este hueso [un fémur] una longitud superior a los doce metros y el volumen de un elemento de dos metros y pico de envergadura». Como podía verse, se concedía a Cuvier una gran parte de la fama del descubrimiento del Megalosaurus y, con ello, de los dinosaurios. Con su inspirada obra sobre anatomía comparada y la extinción de las especies, Cuvier pasó a ser al menos el padrino intelectual de los descubrimientos realizados en la década de 1820 a 1830. En contraste con el Iguanodon vegetariano de Mantell, los dientes en forma de sierra del espécimen de Buckland eran los propios de un carnívoro. Pero Buckland advirtió otras cosas en aquellos dientes. Estaban embutidos en unos alvéolos, como ocurre con los dientes de los cocodrilos, mientras que los dientes de los lagártidos están directamente unidos al maxilar. Después de que Buckland citara algunas razones anatómicas que le llevaban a creer que aquel espécimen no era simplemente un cocodrilo antiguo, no fue capaz de seguir adelante con el tema y nunca se dio cuenta de la importancia de ese rasgo. Como más tarde escribiría Edwin H. Colbert, una moderna autoridad en dinosaurios, eso significaba una «prueba irrefutable» de que el Megalosaurus no era un lagarto gigante, como parecieron asumir Buckland y Cuvier, sino «algo nuevo, un reptil de características hasta entonces nunca imaginadas». Casi todo lo relacionado con la historia de la Tierra, permanecía ignorado y parecía inconcebible a comienzos del siglo XIX. Hacía ya mucho tiempo que la ciencia había desechado la idea precopernicana de un pequeño universo cuyo centro era la Tierra, con lo que se liberaba al espíritu humano para que pudiera pensar libremente sobre las dimensiones virtualmente ilimitadas del firmamento.
En aquellos días nadie conocía la edad del planeta Tierra, unos 4.600 millones de años, ni podía imaginar la progresión de la vida que aparentemente había comenzado a finales de los primeros mil millones de años, quizá hace ya 4 200 millones de años. Al principio el planeta era estéril. Las tierras secas adquirieron formas marcadas notablemente por los cráteres producidos por los meteoritos; estaba iluminado por los fuegos de innumerables volcanes y drenado por la lluvia que alimentaba con aguas frescas a los recientes océanos. El aire estaba formado de nitrógeno, dióxido de carbono y otros gases, pero aún no contenía oxígeno. Había grandes reservas de compuestos con carbono e hidrógeno y algunos otros escasos productos químicos se habían acumulado en la tierra y en las aguas. Las primeras formas de vida, las algas y bacterias que proliferaron, aprendieron a usar la luz como fuente de energía para el crecimiento. Los organismos fotosintéticos que siguieron descomponían el agua para capturar el hidrógeno con lo cual dejaban libre el oxígeno para constituir el medio ambiental. El aire comenzó a contaminarse con el oxígeno, lo cual pudo haber llevado a la destrucción de muchas antiguas especies. La extinción formaba ya parte del proceso de la vida en la Tierra casi en la misma medida que lo sería posteriormente. De esa primera catástrofe global surgieron nuevas formas de vida que utilizaban el oxígeno. La atmósfera, enriquecida con oxígeno, resultaba mucho mejor conductora de los procesos químicos mucho más complejos que habían creado las primeras formas de vida animal. Los animales de esqueleto externo -los trilobites, los corales y los escorpiones de mar- florecieron hace quinientos noventa millones de años. En un espacio de pocos millones de años, en una explosión de nueva vida sin precedentes, hicieron su aparición en la Tierra casi todos los grandes grupos de invertebrados con algunas partes duras en sus cuerpos. Después llegaron los peces, los primeros animales con espina dorsal. Sin embargo, la vida seguía confinada en el interior de las aguas. Pero cuando se alzaron las tierras y surgieron fuera de las aguas y la vida vegetal comenzó a crecer en sus orillas, algunos peces con pulmones salieron fuera del agua apoyándose en sus aletas y establecieron, por decirlo así, una cabeza de puente hace aproximadamente unos cuatrocientos millones de años. Estas especies se desarrollaron en especies anfibias, los antepasados de las ranas, sapos y salamandras actuales. La conquista de las tierras secas por la vida se completó hace unos trescientos millones de años con la presencia de reptiles, los primeros vertebrados independientes del medio acuático.
De esos primeros reptiles descienden casitodos los animales vertebrados que viven en la Tierra en la actualidad, incluidos los seres humanos, así como muchos otros animales desaparecidos, especialmente los dinosaurios. Pero a principios del siglo XIX la mayor parte de los científicos no podían tener una idea del tiempo a escala tan amplia. Podían aceptar la teoría de la extinción de Cuvier, como un primer paso hacia esa comprensión. Pero no podían concebir a los dinosaurios y todo su mundo, porque, en cierto sentido, para ello antes tenían que descubrir el tiempo. Cuvier, por ejemplo, no pudo hacer otra cosa más que sugerir una edad de «varios miles de siglos» para los fósiles contenidos en las capas de tierra de los alrededores de París. Y eso pese a ser un científico en la tradición de la Era de la Ilustración, abierto a las nuevas ideas derivadas de la investigación científica. Muchos otros científicos, sobre todo en Gran Bretaña, se aferraron a determinadas presunciones que militaban contra toda comprensión de la vida en una amplia escala en el tiempo. Partían de la presunción de que Dios había creado la Tierra en un período de tiempo relativamente corto. Debido a ello no había tiempo suficiente para que la naturaleza pudiera tener un pasado que difiriera tanto del presente. Aceptaban la tesis de la inmutabilidad de las especies, por lo que la vida en el pasado no podía ser tan distinta de la de nuestros días. Creían que el mundo había sido creado al servicio y en beneficio del hombre, y de aquí las dificultades que tenían en concebir el mundo sin el hombre, un mundo dominado por especies no humanas.Aunque las pruebas demostrando lo contrario estaban por todas partes y comenzaban a llamar la atención, una gran parte de la Europa cristiana seguía dispuesta a creer que la Tierra no podía tener más de seis mil años. Esta barrera en lo temporal, levantada en nombre de la religión, parecía insuperable. Los ingleses, que hicieron posiblemente los primeros descubrimientos del dinosaurio, e incluso el gran Darwin, crecieron con el convencimiento de que Dios había creado la Tierra y al hombre, así como a todas las demás especies animales, en el año 4004 antes de Cristo. Cuando el botánico sueco Cari von Linneo dijo que «le gustaría creer» que la Tierra era más vieja, no obstante, cumpliendo lo que creía un deber apartó la cuestión afirmando que «las Escrituras no podían permitir una cosa así». Sin embargo, otros fueron menos precavidos. El naturalista francés Georges Louis Leclerc, conde de Buffon, escribiría a mediados del siglo XVIII: «El mejor trabajador de la naturaleza es el tiempo» y para él seis mil años apenas si resultaban suficientes para justificar la existencia de sedimentos y fósiles a tales profundidades en todo el planeta. En su cálculo incluía una estimación basada en el tiempo que necesitaría para solidificarse una esfera en estado de fusión del tamaño de la Tierra. Según eso la edad de la Tierra tenía que ser de al menos 75.000 años y posiblemente hasta medio millón de años. En el mismo siglo Immanuel Kant, en suCosmogonía, le calculaba al mundo una existencia de varios millones de años.

En la época de Mantell y Buckland, las pruebas de la existencia de una vida antigua extinguida y de las «catástrofes» que dieron nuevas formas a la superficie terrestre, exigían nuevas interpretaciones de la edad de nuestro planeta. La descripción de Cuvier de las extinciones asociadas con catástrofes periódicas, resultaba atrayente para muchos científicos como Buckland porque ofrecía la posibilidad de reconciliar la geología con el Génesis. La llamada teoría catastrófica añadía años incontables a la edad de la Tierra. Después de cada catástrofe, según los inspirados discípulos de Cuvier en la Gran Bretaña, Dios comenzaba una vez más con una nueva creación. El libro del Génesis trataba tan sólo de la última de esas creaciones. El diluvio universal, con el arca de Noé, era, presumiblemente, la última de esas catástrofes y no de las que produjeron las aniquilaciones clásicas, porque de acuerdo con la Biblia fueron varias las parejas de criaturas que lograron escapar a la extinción producida por el diluvio. Cuando Darwin zarpó a bordo del Beagle en 1831, él mismo, al igual que la mayor parte de los ingleses, no ponía en duda la verdad literal de la Biblia. El capitán Robert FitzRoy, también un educado caballero, estaba convencido de que el viaje le daría a Darwin la oportunidad de encontrar pruebas de cómo se produjo la aparición de las primeras criaturas en la Tierra y que esto se produciría de acuerdo con el Génesis. Darwin y FitzRoy eran claros representantes de su época. Durante el viaje, Darwin empezó a dejar de lado su antiguo bagaje intelectual. Leyó un nuevo libro de geología, del que era autor Lyell. De esa lectura Darwin obtuvo una nueva perspectiva en el fondo del tema de la posible edad y de la historia de la Tierra. La geología le facilitaba esta nueva perspectiva en el tiempo que estaba promovida por las ideas de Lyell sobre la naturaleza larga, lenta y cíclica de la historia de la Tierra, en contraste con la teoría del catastrofismo de Cuvier y Buckland y mediante la naciente posibilidad de reconstruir el pasado por medio de las reliquias de una vida anterior desaparecida, los fósiles. El descubrimiento del tiempo se fue haciendo por medio de una cuidadosa y atenta lectura de la Tierra y los fósiles en ella contenidos. Rompió la barrera del tiempo en las últimas décadas del siglo XVIII. Aunque sus logros permanecieron largo tiempo sin ser debidamente reconocidos, hasta después de 1830, llegó a tiempo para ejercer una importante influencia en el descubrimiento de los dinosaurios y en la teoría de la evolución de Darwin. James Hutton, geólogo, médico, naturalista, químico escocés del siglo XVIII, no encontró pruebas de la existencia del diluvio universal. Por el contrario, creía que toda formación rocosa, con independencia de su antigüedad» parecía derivarse de las rocas más antiguas. Para Hutton todas las características del paisaje actual eran producto de un cambio gradual. «Desde la cima de las montañas a las orillas del mar -declaró-, todo se halla en estado de cambio».
En un mensaje dirigido a la Royal Society de Edimburgo, en 1785, y en el libro Theory of the Earth, publicado en 1795, Hutton describió la Tierra como una «máquina mundial», dinámica y autorregulada, gobernada por leyes naturales. No había necesidad de invocar acontecimientos singulares de creación o revolución para explicar la faz de la Tierra. Dado un tiempo suficiente, la actividad geológica en sus presentes niveles podía haber producido todas las rocas y determinado la actual forma de la superficie de la Tierra. De acuerdo con las ideas de Hutton la superficie de la Tierra se va deteriorando y renovando al mismo tiempo, por el ciclo de cuatro etapas: erosión, deposición, consolidación y elevación. Los materiales erosionados eventualmente se depositaban como sedimentos en el fondo de los mares y océanos en la segunda fase del ciclo. Allí, los sedimentos se hacían compactos y se consolidaban en capas de rocas, en la tercera etapa. Todo esto había sido reconocido por algunos de los predecesores de Hutton. Quedaba una cuarta fase en el ciclo, advirtió Hutton, pues de no ser así los continentes se irían alisando y los océanos acabarían por llenarse. Tenía que haber una fuerza restauradora y vio en los volcanes el mecanismo por el que actuaba la cuarta fase del ciclo. Hutton vio en el vulcanismo la manifestación del poder «licuador y de la fuerza expansiva del fuego subterráneo». Los volcanes en erupción lanzaban nuevas rocas, basalto y granito procedentes de las profundidades de la Tierra. Se creía que el granito era la más antigua de todas las rocas, que se habían creado como sedimentos en el fondo del océano universal. Hutton, que había visto en las tierras altas de Escocia muchas vetas de granito que atravesaban capas más antiguas de roca sedimentaria, las rocas areniscas y la pizarra, pensaba de manera distinta. El calor interior de la Tierra no sólo suministraba nuevas rocas, postularía más tarde Hutton, sino que era también responsable de que las rocas sobresalieran fuera del mar y se plegaran para formar nuevas montañas y continentes. Vio la prueba de ello en las angulosas irregularidades que surgían en los lados de las colinas de Escocia. Esos estratos, plegados y torturados, implicaban un proceso de elevación de la corteza terrestre «que tenía por origen el calor subterráneo». Los fósiles marinos descubrían el origen de la capa de tierra. John Playfair, el biógrafo de Hutton, viendo una de esas reveladoras deformaciones en un viaje en compañía de Hutton por ese terreno, escribió: «Cualquiera hubiese visto allí la clara evidencia de las distintas formaciones de esas rocas y de los largos intervalos que separaban entre sí esas formaciones, ¿las habíamos visto surgir desde el fondo de las profundidades? La mente parecía aturdirse cada vez más al contemplar tan profundamente los abismos del tiempo».
La geología ortodoxa se dirigió en dirección al catastrofismo de Cuvier, que no requería una cantidad de tiempo indefinida y, más aún, que invocaba la existencia de océanos primitivos que presentaban un notable parecido con la inundación producida por el diluvio universal. Se dejó en manos de Charles Lyell, nacido el año 1797, el mismo año en que murió Hutton, el establecer y afirmar la teoría de Hutton como la nueva ortodoxia geológica. Esto lo consiguió con una obra en tres volúmenes titulada Principies of Geology. Fue el primero de esos volúmenes el que Darwin se llevó consigo durante su viaje en el Beagle. El concepto de Lyell sobre el cambio gradual en el transcurso de un tiempo ilimitado, preparó el camino para la geología y la paleontología modernas. Influyó en Darwin y lo llevó a considerar el tiempo a una escala suficiente para el lento proceso de la evolución y concedió a los paleontólogos la necesaria libertad para imaginarse mundos desaparecidos, habitados por criaturas que vivieron y se extinguieron mucho antes de que nacieran los patriarcas bíblicos. Los cataclismos han modificado muchas veces la faz de la Tierra a lo largo de los tiempos. La dinámica del suelo de los fondos marinos extendiéndose y la elevación que hizo surgir a los continentes, causan las convulsiones tectónicas de las placas. Esto y las catastróficas inundaciones de Cuvier, constituyen procesos continuados cuyo efecto es acumulativo sobre el clima y la vida. El geólogo inglés William Smith sentía curiosidad por los fósiles y aún más sobre la forma como algunas capas de roca contenían fósiles siempre situados en un mismo orden determinado. A Smith se le ocurrió la idea de que los estratos rocosos podían ser identificados por los fósiles característicos que había en su interior. Smith había reconocido el segundo de los dos principios fundamentales de la estratografia. Se había establecido ya que los estratos de la Tierra se situaban en un orden regular de modo que, en circunstancias normales, sin trastornos excepcionales, los estratos inferiores son las capas más antiguas. Esto le sirvió a Smith como principio conductor para su cartografía geológica, pionera en Inglaterra. Su nuevo descubrimiento fue que los restos orgánicos enterrados en las capas geológicas se sucedían en el mismo orden de regularidad. Para Smith era importante que el aspecto más distintivo de una capa particular de tierra no era su grosor o composición mineral, puesto que éstas variaban según el lugar en el mismo estrato, dependiendo del origen de los sedimentos, sino los fósiles que había en su interior. Ciertos fósiles que Smith encontró en un estrato, nunca los halló en ningún otro. Eran los restos de criaturas que habían vivido en el tiempo en que se estaba depositando un determinado estrato y no en ningún otro. Éstos son llamados en la actualidad «fósiles índices».
Observando «el orden y la regularidad maravillosos»de esos fósiles índice, Smith afirmó que era posible establecer una correlación entre los estratos en pliegues terrestres separados entre sí por muchos kilómetros, incluso en distintos continentes. Los fósiles más humildes son los más útiles en este aspecto. Dinosaurios, Mosasaurus y tigres prehistóricos eran demasiado grandes y raros para servir de guía a los geólogos que trataran de identificar un estrato. Dado que los estratos, por muy distintos que puedan parecer, son de la misma edad cuando contienen los mismos fósiles índice, William Smith había descubierto un medio para medir el tiempo. No el tiempo absoluto en miles y millones de años, puesto que esas mediciones tenían que esperar aún un siglo, sino el tiempo relativo. Según Smith, cada capa de roca representaba un intervalo de tiempo identificable por un determinado conjunto de fósiles y la secuencia de las distintas capas representaba la progresión del tiempo en el acontecer geológico. De esta manera, la vida en sí, la vida pasada preservada en forma fósil, pasó a ser la clave para la comunicación del tiempo geológico. Las posibilidades que tiene un organismo de convertirse en fósil son mayores si posee un esqueleto resistente o una concha mineralizada. La circunstancia de que se produzca un enterramiento rápido, repentino y total, aumenta las posibilidades de petrificación. El enterramiento protege a los organismos contra el oxígeno y otros agentes de la corrupción. Esto facilita que transcurra el tiempo suficiente para que se produzca la mineralización. El agua que contiene minerales disueltos penetra en los poros del hueso o la concha y, lentamente, destruye mucho de su materia original reemplazándola, molécula a molécula, por sus minerales. El resultado es un fósil pétreo de la forma original, que contiene bien poco, o nada, de la materia orgánica original. Las condiciones más favorables para que se produzca esa fosilización existen en los sedimentos del mar, en las ciénagas, en los pantanos y en las pequeñas charcas de agua estancada. Los animales que mueren en suelo seco tienen menos posibilidades de fosilización, un hecho que tiende a distorsionar el registro de los fósiles. Consecuentemente los científicos nunca llegarán a saber cuántas especies de dinosaurios vivieron a la sombra delIguanodon de Mantell o del Megalosaurus de Buckland y que se extinguieron sin dejar tras ellos fósiles que recordaran su existencia. Cada edad de la Tierra dejó tras de sí un índice incompleto de fósiles. De los tres millones de especies de seres vivos que existen en el mundo en la actualidad, sólo una pequeña fracción tiene una posibilidad de dejar tras sí fósiles identificables. El más extendido de los órdenes animales de la Tierra comprende trescientas mil especies de escarabajos, un hecho que ha llevado al especialista en genética J. B. S. Haldane a responder, cuando se le preguntó cuáles eran los rasgos característicos del Creador que se habían puesto en evidencia en la vida de la Tierra: «Debió de tener un especial cariño por los escarabajos».
Pero serán muy pocos los que sobrevivirán como fósiles, si es que lo hace alguno. La mayor parte de las restantes especies de seres vivos en la actualidad son plantas, es decir, pobres candidatos a la fosilización. Existen 8 600 especies de aves y 4 000 de mamíferos, cuyas posibilidades son algo mejores. Sin embargo, sólo pocas de esas especies dejarán fósiles que puedan sugerir qué tipo de vida existía en la época en la cual una de las especies existentes en particular empezó a comprender el significado y la importancia de los fósiles. Esta comprensión de los fósiles comenzó con dos científicos del siglo XVII que publicaron el primer análisis del fenómeno. Fueron Robert Hooke y Niels Stensen, este último más conocido por la latinización de su nombre, Nicolaus Stenonis, o simplemente Steno. Hicieron observaciones avanzadas a las que se adhirieron pocas personas hasta un período muy posterior. Hooke, un matemático inglés con gran prestigio intelectual en el tiempo de Isaac Newton y Edmond Halley, ofreció las pruebas de que los fósiles eran los restos de organismos convertidos en piedra o las impresiones dejadas por ellos. Su razonamiento le hizo sobrepasar la línea de la sabiduría convencional. Dado que son tantos los fósiles que tienen origen marino, dijo, indican que la superficie de la Tierra ha sufrido una marcada transformación desde la creación, en la cual el mar cedió el paso a la tierra seca, después el mar cedió paso a la tierra, y así una y otra vez, como Cuvier, con pruebas más efectivas, establecería más tarde. Con respecto al significado de los fósiles, Hooke escribió: “Comprendo humildemente (aunque es posible que haya quienes piensen que se le concede demasiada importancia a algo tan trivial como una concha putrefacta) que los hombres actúan por lo general con demasiada superficialidad y pasan sin conceder importancia a estas marcas de la antigüedad que la naturaleza nos dejó tras sí como monumentos y caracteres jeroglíficos de transacciones precedentes semejantes a cambios en el cuerpo de la Tierra, que son infinitamente más evidentes y probatorias que cualquier otra cosa que la antigüedad pueda dejar tras sí, como monedas o medallas o cualquier otra cosa hasta ahora conocida, dado que la mayor parte de esos caminos pueden ser falsificados o hechos con arte y destreza… Las conchas fósiles no pueden ser falsificadas ni por la mayor destreza que en el mundo pueda darse, ni nadie que examine de manera imparcial sus apariencias puede dudar de que son realmente lo que parecen. Y aunque hay que aceptar que resulta muy difícil leerlas y trazar su geocronología y fijar los intervalos de tiempo en los cuales ocurrieron tales y cuales catástrofes o mutaciones. Sí, esto no es imposible“.La referencia de Hooke a las medallas y monedas delataba, desde luego, su creencia de que los fósiles se corresponderían con el período de la vida humana.
Steno era un médico danés que, mientras servía a un duque florentino, se tomó el tiempo suficiente para investigar los fósiles marinos de la Toscana italiana. Reconoció su estructura interna debido a sus conocimientos de anatomía y demostró que los fósiles sólo podían ser reliquias de criaturas vivas. Pero Steno acabaría por convertirse al catolicismo y llegaría a obispo, por loque no podía permitirse imaginar que los fósiles fuesen más antiguos que el diluvio universal. Tampoco podía creer que los huesos de elefante, que estaban siendo descubiertos en aquellos días, fuesen prehistóricos. Eran, así lo declaró, los restos de los elefantes africanos que Aníbal trasladó desde África a Italia. Sin embargo, Steno supo hacerse una perspectiva más acertada que ninguna otra en el siglo XVII sobre la conformación de la Tierra en capas o estratos. Llegó a la conclusión de que esos estratos se habían ido asentando en forma de sedimentos acuosos que más tarde se solidificaron para transformarse posteriormente en dura roca, en que los estratos superiores eran más jóvenes que los inferiores. El principio de superposición es en la actualidad un axioma de la estratografía. Fue el principio aplicado por Cuvier, cuando reconoció el hecho de la extinción de las especies a lo largo del pasado y postuló que eso ocurrió como consecuencia de una serie de catástrofes ampliamente extendidas. Al igual que Hooke había sabido ver el significado de los fósiles para determinar el tiempo geológico, aunque quedándose por detrás de las observaciones más precisas de Smith, Steno reconoció que los estratos de la corteza terrestre contenían en sí un registro cronológico de los más importantes acontecimientos ocurridos en la Tierra en el pasado y que cada uno de los estratos podía ser considerado como un capítulo en la historia de la Tierra. Todos ellos llegaron a aproximarse al descubrimiento de la insospechada edad de la Tierra, pero no profundizaron en ello. Este tema sufrió una detención que duró varias generaciones, hasta llegar a William Smith. Este geólogo alemán aplicó el principio de superposición del estrato de Steno y llegó a establecer con gran certeza el hecho de que diferentes capas de rocas representaban episodios del tiempo geológico, el primer principio de la estratografía. Otros científicos habían identificado y clasificado muchos fósiles para llegar a un consenso en la aceptación de que se trataba de restos de una vida pasada. Pero nadie entre ellos comprendió la relevante relación existente entre los estratos y los fósiles hasta que Smith puso juntas ambas cosas y mostró cómo se debía leer el tiempo geológico. Durante las siguientes décadas, a medida que la secuencia de las capas de la corteza terrestre y los fósiles se hizo más familiar, los científicos empezaron a diseñar la escala temporal geológica que está en uso en nuestros días, que se convertiría en el calendario general de la geocronología (ver artículo “Eras geológicas“.

Las estaciones eran las eras: precámbrica, paleozoica, mesozoica y cenozoica. Las divisiones, con excepción de la precambriana, fueron propuestas en la década 1840-1850 por John Phillips, sobrino y aprendiz de William Smith. Reflejaban las claras transiciones de un período a otro, subrayadas por extinciones masivas, por el paso de un sistema de vida predominante a otro, tales como la vida primitiva del paleozoico, los grandes reptiles del mesozoico y los mamíferos del cenozoico. Anteriormente, en la era precámbrica, que abarcaba el período que va desde la formación de la Tierra hasta la aparición de vida multicelular abundante, los estratos contenian un número muy escaso de fósiles. Los meses del tiempo geológico son las divisiones que ahora conocemos como períodos. Son, siguiendo el orden de antigüedad: cámbrico, ordovícico, silúrico, devónico, carbonífero y pérmico, todas ellas divisiones de la era paleozoica. En los Estados Unidos las formaciones carboníferas se clasifican como los períodos mississípico y pensilvánico. Por lo general, los períodos llevan los nombres de las zonas en los que fueron estudiados detalladamente por vez primera. Los períodos ordovícico y silúrico, por ejemplo, llevan los nombres de antiguas tribus galesas. Cámbrico deriva del nombre romano de Gales; pérmico, de la provincia de Perm, en Rusia; devónico, de Devonshire. El carbonífero es un nombre claramente descriptivo, el «período del carbón», o los estratos en los cuales se encuentra carbón. Para los cazadores de dinosaurios los tres períodos más importantes son los del mesozoico: triásico, jurásico y cretáceo. Toman sus nombres de los estratos del sur de Alemania, con una estructura claramente tripartida de sus rocas; de las montañas del Jura y de la palabra latina que significa cal o greda, debido a La composición calcárea de los estratos en las cercanías de París que sirvieron para definir y categorizar al período. El cenozoico está dividido en dos períodos principales: terciario y cuaternario. Debido a la gran abundancia de fósiles hallados, esos dos períodos pasaron a ser subdivididos en épocas conocidas por paleoceno, eoceno, oligoceno, mioceno, plioceno, pleistoceno, la época de la más reciente de las eras glaciales, y holoceno, que incluye los últimos diez mil años hasta el momento presente. Sin embargo, éste era un calendario sin fechas. Dado que los geólogos no podían asignar unas edades absolutas a los estratos, los paleontólogos que hicieron los más importantes descubrimientos relacionados con el dinosaurio en el siglo XIX y ya entrado el XX sólo sabían que esas criaturas vivieron en uno u otro de los períodos del mesozoico. Podían decir, por ejemplo, que los pequeños coelosaurios procedían de rocas que indicaban que habían vivido en el triásico. Igualmente pudieron asignar los Aliosaurus, Brontosaurus y Stegosaurus al jurásico. ElIguanodon y el Tyrannosaurus eran definitivamente criaturas del cretáceo.
Así, los paleontólogos saben cómo situar los distintos dinosaurios en la sucesión del tiempo, pero se encontraban absolutamente perdidos a la hora de determinar las edades absolutas de esos períodos en los que predominaron los dinosaurios. Es lo mismo que si los historiadores conocieran sólo edades relativas y tuvieran que limitarse a situar el descubrimiento de América en un período sin fecha concreta situado entre las cruzadas y la revolución industrial. Los científicos que trataron de calcular la edad absoluta de la Tierra se presentaron con distintas respuestas, ninguna de ellas satisfactoria. Los cálculos efectuados ni siquiera se aproximaban a la verdadera edad del planeta, pero nadie podía estar seguro de ello hasta que los científicos se dieron cuenta de que las rocas contenían en sí su propio «reloj» atómico que estaba en marcha desde el momento mismo de su formación. En 1905, Bertram Boltwood, un químico de la Universidad de Yale, observó una relación entre el uranio y el plomo. Los átomos del mismo elemento, con diferentes pesos atómicos, debido a las variaciones en el número de electrones, se llaman isótopos. La decadencia radiactiva es un proceso espontáneo en el cual un isótopo de un elemento madre pierde partículas de su núcleo para formar un isótopo de un nuevo elemento hijo. Algunos isótopos que se degeneran lentamente son utilizados como relojes atómicos. La degeneración del uranio, un fenómeno recientemente descubierto, produce plomo. La cantidad de plomo mezclada con el uranio ofrecería una medida del tiempo transcurrido desde la formación del mineral. Muestras de rocas de las antiguas formaciones, contenían una proporción menor de uranio en relación con el plomo que aquellas otras formaciones posteriores, es decir, más jóvenes. Formaciones de la misma edad fósil mantenían la misma relación plomo-uranio. Otro reloj atómico que en la actualidad es de utilidad para los geólogos se basa en la relación de los isótopos de torio y plomo, de rubidium y estroncio, y del potasio con el argón. Cuando Emest Rutheford, un físico nacido en Nueva Zelanda, calculó el ritmo del cambio del uranio al plomo, los científicos comenzaron con sus primeros intentos de obtener la edad radiométrica de la roca y de la propia Tierra. La edad calculada por Boltwood ascendía hasta los dos mil millones de años. Posteriores mejoras y perfeccionamientos en el método, conjuntamente con la creación de instrumentos analíticos más sofisticados, produjeron un aumento progresivo de la edad calculada para la Tierra. Un libro sobre geología publicado poco antes de la segunda guerra mundial informó de una edad de tres mil millones de años. Un texto de los años 1950-1960 habló de los 4 500 millones de años. En la actualidad, con una certeza que nunca fue posible con anterioridad, la edad fijada es de 4600 millones de años. Los «relojes atómicos» en las rocas, permitieron a los científicos asignar fechas absolutas al calendario del tiempo geológico: 590 millones de años para el comienzo del mesozoico; y 65 millones para el final del cretáceo y de los dinosaurios. James Hutton descubrió la magnitud del tiempo geológico y William Smith aprendió a leer los estratos y los fósiles.
Al Megalosaurus de Buckland y al Iguanodon y el Hylaeosaurus de Mantell, siguieron nuevos descubrimientos. Para 1841, a esa serie de reptiles arcaicos se unieron el Macrodontonphion, elThecodontosaurus, el Paleosaurus, el Plateosaurus, el Cladeidon y el Cetiosaurus y sus restos fósiles trataban de decir a los científicos algo sobre la vida en el mesozoico. Esas criaturas recién descubiertas parecían ser lo suficientemente distintas de los reptiles modernos o de los gigantes marinos, como el Mosasaurus de Cuvier o el Ichthyosaurus de Mary Anning, como para merecerse un nombre familiar para ellos solos. Y así pasaron a ser conocidos colectivamente por el nombre más fácil de pronunciar de Dinosauria, es decir, de dinosaurios. Ese nombre se debe a la inspiración de Richard Owen, un brillante especialista en anatomía y paleontología, a quien en su tiempo se le llamó el Cuvier inglés. Su ascensión hasta tan altas cumbres profesionales fue muy rápido. Fue él quien dio nombre a los dos fósiles descubiertos más recientemente: elCladeidon, del triásico, y el Cetiosaurus, del jurásico. En la reunión anual de la Asociación Británica para el Progreso de las Ciencias, celebrada en Plymouth el 2 de agosto de 1841. Owen expresó muchos reparos con respecto a los reptiles fósiles, en particular sobre elIguanodon, el Megalosauros y el Hylaeosaurus. Eran, indudablemente, criaturas gigantescas pero, ¿se trataba de lagartos gigantes? Aunque ya Mantell había conjeturado que eran algo distintos de los otros reptiles arcaicos, eran muchos los científicos que los consideraban simplemente como lagartos excesivamente desarrollados. En su escrutinio de los fósiles, Owen observó muchas características que, así se lo dijo a su audiencia en Plymouth, le «ofrecían suficientes motivos para establecer una tribu o suborden distinto de reptiles saurios» para los cuales propuso el nombre dinosaurios {Dinosaurio). Owen acuñó ese nombre a partir del griegodeinos, que significa «terrible» o «espantosamente grande» y sauros que significa lagarto o reptil. Dinosaurio, el terrible lagarto. Owen estableció sus razones para una nueva clasificación de la vida animal extinguida. Su principal argumento era que, a diferencia de otros saurios gigantes, aquéllos eran criaturas terrestres más que acuáticas. Otra diferencia era que, en contraste con otros reptiles, tenían la característica común de poseer cinco vértebras fundidas en el cinturón pélvico. Sus enormes esqueletos, ya que aún no habían sido descubiertos los pequeños dinosaurios, sugerían que tenían cuerpos más masivos y pesados que los alargados lagartos y mosasaurios. A Owen le recordaban a los mamíferos paquidermos. «De la forma y el tamaño de las costillas -dijo- se deduce que su tronco era más ancho y grueso en sus proporciones que el de los saurios modernos y, sin duda, se alzaba del suelo apoyado en extremidades proporcionalmente más anchas y en especial más largas, de modo que el aspecto general del Megalosaurus en vida debía de parecerse proporcionalmente a los grandes mamíferos cuadrúpedos que ahora pueblan la Tierra, y que debieron de ocupar en la era olítica (los tiempos antiguos cuando ciertas rocas, como la piedra caliza, se sedimentaron) el lugar que hasta entonces tenían los grandes reptiles del extinguido orden de los dinosaurios»

Owen propuso que los cálculos de sus dimensiones se basaran en el tamaño y el número de las vértebras, un método que aún se sigue empleando en la actualidad. Eso nos da una longitud para el Iguanodon de ocho metros, y de nueve metros para el Megalosaurus. Aunque creía que las especies habían cambiado con el paso del tiempo, Owen estaba convencido de que cada uno de los animales dentro de uno de los grandes grupos eran variaciones de un mismo tema, del «arquetipo ideal» y de que la «mente divina que había planeado el arquetipo también conocía anticipadamente todas sus modificaciones». Su evolución planeada era antitética a las ideas de Jean Baptiste Lamarck, contemporáneo de Cuvier. Lamarck creía en la evolución, pero no en la extinción; las especies no desaparecían sino que iban cambiando gradualmente, transformándose en algo diferente y, a su juicio, mejor. Aunque Darwin aún tenía que publicar su teoría, la evolución era ya materia discutida en el campo científico. Aquellos que seguían creyendo que Dios había creado las especies en sus formas presentes e invariables, o los que, como Owen, creían que Dios «preconocía» todas las modificaciones, dirigieron el ataque contra el esquema de Lamarck sobre las cosas. En 1809 Lamarck había postulado la existencia de una tendencia intrínseca de todos los organismos hacia la perfección, por lo que se desarrollaban progresivamente desde las formas más simples hacia las más complejas. Ésta era la tendencia «observada» en la naturaleza. Pero no era ésa la forma como Owen la veía cuando contempló los fósiles de lo que él llamaría dinosaurios. Si de veras existía como afirmaba Lamarck un ascenso de la simplicidad a la complejidad, argüía Owen, los principales reptiles que en la actualidad poblaban el mundo no serían unas criaturas comparativamente tan pequeñas. Los fósiles del Iguanodon y del Megalosaurus, afirmaba, revelaban que los dinosaurios eran «la cumbre de la creación reptiliana» y la «más cercana aproximación a los mamíferos». Pero, ¿dónde estaban los dinosaurios? Muertos hacía ya mucho tiempo, y todos y cada uno de los tipos de reptiles que los habían sucedido eran criaturas mucho más simples y menos evolucionadas que ellos. Si esto era evolución, se trataba de una evolución retrógrada. Owen estaba exponiendo un punto débil en la teoría de Lamarck, en su concepto de la evolución. Con satisfacción, porque sabía la importancia de su argumento, Owen subrayó enfáticamente la superioridad de los dinosaurios sobre otros reptiles. Adrián J. Desmond, en The Hot-Blooded Dinosaurs, escribió: «Literalmente Owen pobló su tierra mesozoica con dinosaurios como una estrategia contra los evolucionistas».
Owen siguió adelante y declaró que los dinosaurios no se habían desarrollado a partir de un orden más bajo de reptiles, sino que habían sido creados por Dios. Más aún, Dios los había colocado en la Tierra en un tiempo particular del mesozoico porque reinaban condiciones especiales que les eran muy favorables. Debido a la abundancia de grandes reptiles y a la ausencia de todo mamífero de importancia, Owen llegó a la conclusión de que la atmósfera mesozoica era deficiente en oxígeno. Razonó que los reptiles, al ser animales de sangre fría y menos energéticos, requerían menos oxígeno que los mamíferos de sangre caliente. Posiblemente, una «vigorización» de la atmósfera, con más oxígeno, hizo la Tierra inhabitable para los dinosaurios. Ésta parece ser la primera teoría para explicar la extinción de los dinosaurios, aunque, ciertamente, no habría de ser la última. Hacia el final de su informe, Owen introdujo algunas ideas provocativas sobre el metabolismo de los dinosaurios. Los dinosaurios, dijo, es posible que tuvieran una vida más vigorosa que la que se asocia con los reptiles. Su «estructura torácica» indicaba que debieron de tener un corazón con cuatro cámaras, más parecido al de los mamíferos y las aves que al corazón con tres cámaras de reptiles. «Su superior adaptación a la vida terrestre», dijo Owen, era una prueba más de que tenían que «disfrutar de la función de un órgano central de la circulación altamente organizado en un grado bastante aproximado al que ahora caracteriza a los vertebrados de sangre caliente». Richard Owen había presentado el primer informe inteligible y generalizado sobre los dinosaurios. Posiblemente sólo un hombre de su arrogancia se hubiera atrevido a sacar tan amplias deducciones de tan pocos huesos, que representaban los restos parciales de no más de nueve géneros de reptiles arcaicos. Sin embargo, cualesquiera que fueran sus motivos, pese a la debilidad de algunas de sus pruebas, aquel día de 1841, Owen «creó» el dinosaurio. Apropiadamente Owen aisló al dinosaurio y lo separó de la tipificación usual de los reptiles. Pero casi nada de lo que se sabe sobre ellos parece indicar que constituyan un grupo natural único. Desde el pasado siglo XIX, por el contrario, la mayor parte de los científicos han llegado a la conclusión de que los llamados dinosaurios consisten en dos grupos separados que estaban emparentados muy lejanamente. Estos grupos eran los saurischias y los ornitichias. Como los nombres sugieren, ambos grupos tenían anatomías claramente diferenciadas que posiblemente los hacían no estar más relacionados entre sí de lo que puedan estarlo con otros miembros del árbol genealógico de los reptiles, como los cocodrilos o los pterosaurios. Conjuntamente, los saurischias y los ornitichias, los crocodílidos y los pterosaurios son miembros de un amplio conjunto conocido como archisaurios. Estos cuatro órdenes se desarrollaron a partir de los más primitivos de ellos, los tecodontes. De los archisaurios, sólo algunos crocodílidos -los aligátores y los cocodrilos del Ganges, así como los auténticos cocodrilos- han sobrevivido hasta la actualidad. Los archisaurios son descritos diversamente como un superorden o una subclase de los reptiles. El Tyrannosaurus rex era un lagártido (saurischia), bípedo (pterópodo), y un animal carnívoro (camosauria). No hay nada que indique que fuese realmente un dinosaurio.
Los Homo sapiens son (somos) vertebrados que alimentan a sus hijos con leche procedente de las glándulas mamarias de sus hembras (mamíferos), desarrollan a sus hijos in útero tienen manos y pies con cinco dedos, dotados de uñas en vez de garras, como los primates, tienen visión dirigida hacia adelante y gran cerebro, poseen extremidades superiores especializadas para la manipulación y son bípedos para de ese modo poder cazar utilizando armas que sostienen en sus extremidades anteriores.Los dinosaurios, tanto sauriquianos como omitisquianos, comparten una característica que falta en los demás reptiles. Todos se alzan en posición «completamente erecta» con sus patas traseras soportando el peso de cuerpo por detrás. En esto son más parecidos a los mamíferos primates que a los reptiles. La mayor parte de los restantes reptiles son reptadores, con sus extremidades proyectándose a los lados del cuerpo. Los actuales cocodrilos se colocan en una postura semierecta sólo cuando quieren correr trotando, lo cual no es frecuente. La estructura de los miembros que soportaban a los dinosaurios en su postura erecta les permitió, también, a muchos de ellos, desarrollarse en auténticos bípedos, es decir, a mantenerse de pie sobre sus dos patas traseras. En este aspecto eran muy distintos de los restantes reptiles y más parecidos a los primates. Alan Charig, un paleontólogo británico, encuentra en esto «cierta justificación» para el uso del término popular «dinosaurio», para denominar tanto al sauriquiano como al omitisquiano. Un escultor, Benjamín Waterhouse Hawkins trataba de «revivir el mundo antiguo». Trabajando en estrecha colaboración con Owen, Hawkins dio forma con cemento, piedra, ladrillos y hierro a reproducciones de tamaño natural de anfibios, cocodrilos y plesiosáuridos extinguidos, así como a tres dinosaurios a los que dedicó su mayor atención, el Iguanodon yel Megalosaurus Con ello Hawkins creó la falsa ilusión de que el hombre moderno sabía más sobre aquellas bestias de lo que en realidad conocía. Sospechosamente, algunas de las reproducciones parecían ranas y tortugas aumentadas de tamaño. El Iguanodon tenía el aspecto de un rinoceronte reptil, pues Hawkins perpetuó el error de Mantell poniendo un cuerno sobre el hocico de aquella criatura. Se representó al Iguanodon y al Megalosaurus como si fueran animales que anduvieran a cuatro patas, pues los científicos aún no habían descubierto su sorprendente bipedalidad, que se creía patrimonio de los primates. La revista Punch, en un artículo titulado «Diversión en un fósil», ignoraba la naturaleza herbívora del Iguanodon al declarar: «Nos congratulamos de la compañía (de los científicos) en la era en la que vivimos, porque si hubiera sido en un período geológico anterior es muy posible que hubieran ocupado el interior del Iguanodon, pero no para cenar allí».

¿Por qué los dinosaurios fascinan tanto a la humanidad? Uno de los temas principales de la mitología son los héroes Beowulf, Sigfrido o San Jorge luchando contra un dragón. Es el combate contra fuerzas misteriosas. Casi invariablemente el símbolo del terror es un animal enorme, fundamentalmente un reptil. Los dinosaurios tienen tanto parecido con los míticos dragones que tal vez inspiraron su leyenda. Encuentros entre seres prehumanos y dinosaurios en un pasado remoto debieron de dejar cicatrices indelebles en la psique humana. Algunas de las primeras descripciones de los grandes reptiles alimentaron la creencia en el parecido entre dinosaurios y dragones. En 1840, un año antes del informe de Owen, un geólogo británico llamado Thomas Hawkins publicó The Book of Great Sea Dragons, que estaba basado, sin demasiada conexión, en los descubrimientos de Cuvier, Anning, Buckland y Mantell. Las ilustraciones del libro mostraban reptiles monstruosos de la tierra, las aguas y el aire luchando en siniestros paisajes. Esas ilustraciones le recuerdan a un escritor, L. Sprague de Camp, la leyenda de los dragones medievales de tal modo que, al verlas, «uno casi espera ver aparecer en escena a un caballero armado de punta en blanco». Paleontólogos e incluso científicos, analistas de la conducta, parecen encontrarse perdidos a la hora de explicar el porqué de la influencia que los dinosaurios ejercen sobre la psique humana. Debe de ser alguna razón elemental puesto que son los niños los que con más facilidad se sienten subyugados por los dinosaurios y a una edad muy temprana. Los dinosaurios son enormes y terroríficos, alimentan las fantasías. El tamaño y la ferocidad pueden ser un factor de su popularidad porque los dinosaurios de pequeño tamaño apenas si son mencionados. Y todas esas criaturas fueron reales lo que, al parecer, las hace más terribles y excitantes que las criaturas imaginarias de los cómics. Parece evidente que los dinosaurios satisfacen una necesidad humana de misterio y aventuras. En su libro Huntingfor Dinosaurs, publicado en 1969, Zofia Kielan-Jaworowska, una eminente paleontóloga polaca, escribió: “Ningún científico familiarizado con la aventura intelectual de estudiar animales de un pasado remoto, tendrá la menor vacilación en afirmar que el viajar millones de años en el pasado, que es lo que significa el estudio de la paleontología, resulta mucho más fascinante que el más exótico de los viajes geográficos que estamos en condiciones de realizar en la actualidad. El estudio de animales que vivieron en la Tierra hace millones de años no es, simplemente, el estudio de su anatomía, sino en primer lugar, y sobre todo, un estudio del transcurso de la evolución en la Tierra y de las leyes que la gobiernan“.
El estudio de la evolución de animales que habitaron la Tierra durante millones de años y el compararla con la historia de la humanidad -tan breve a escala geológica- coloca la posición del hombre en el mundo de los seres vivos en su auténtica perspectiva y ayuda a contrarrestar las ideas antropocéntricas. De todos los animales prehumanos, las criaturas a las que Owen llamó dinosaurios, y que Hawkins revivió en piedra, se hicieron irresistibles para el público y, también, para un número cada vez mayor de científicos. En noviembre de 1859, veintitrés años después del regreso del Beagle, Charles Darwin hizo pública la teoría que lo inmortalizó y revolucionó la biología y todas las ciencias relacionadas con ella, incluyendo la paleontología. Previendo la controversia que sus ideas habrían de provocar y deseoso de evitar el combate intelectual, Darwin retrasó la publicación de su teoría de la evolución. Posiblemente la hubiera aplazado aún más de no haberse enterado en 1858 de que otro naturalista inglés, Alfred Russell Wallace, había llegado de modo independiente a las mismas conclusiones y estaba dispuesto a publicarlas. Ningún científico, ni siquiera Darwin, que estaba ya a punto de retirarse, podía permanecer impasible y sin hacer nada mientras que otro se apuntaba la gloria de una idea en la cual tenía absoluta prioridad. En vista de ello Darwin se apresuró a publicar, a finales de 1859, su obra maestra On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life. Aunque algunos otros científicos habían especulado con la tradición y el dogma en defensa de la evolución, nadie antes que Darwin y después de Wallace, había concebido una explicación plausible de cómo se desarrollaba esa evolución. Ni siquiera Lamarck, el más destacado de los evolucionistas de principios del siglo XIX. El mecanismo que hacía funcionar la evolución, como creía Darwin, era la selección natural. Ésta fue la idea más profunda y original de Darwin. La selección natural, en oposición a la selección artificial, ya por aquel entonces ampliamente practicada por los criadores de animales y plantas, era cuestión de la «supervivencia de los más aptos», una frase que Darwin no utilizó en la primera edición de su libro, pero que acabaría por caracterizar su teoría. Partiendo de sus experiencias como naturalista del Beagle y de su lectura de los puntos de vista de Thomas Malthus sobre la población, Darwin hizo dos importantes observaciones: que los organismos individuales, dentro de una misma especie, tienen caracteres distintos, y que la mayor parte de los organismos tienen más descendencia de la que logra sobrevivir. Los organismos que adquieran las características que resulten más adaptadas al ambiente sobrevivirán y se reproducirán, dedujo Darwin, y aquellos que no lo hagan morirán. Con el paso del tiempo, y con herencias acumulativas, una especie puede llegar a modificarse hasta el punto de convertirse en algo distinto, una nueva especie diferenciada.
En la metáfora botánica de Darwin, el árbol de la vida crece y cambia con la apariencia de «frescos pimpollos y éstos, si son vigorosos, se ramifican y por todas partes cubren con sus fuertes ramas a las otras más débiles». Ésta es la evolución de las especies por selección natural, de acuerdo con Darwin. Pero ¿dónde se hallaban en los fósiles las formas transicionales entre los ascendientes y los descendientes de los organismos? Darwin, en Origin of Species,reconoce la debilidad en este caso de la selección natural y define a esa ausencia de fósiles de las formas intermedias como «probablemente la más grave y más obvia de las muchas objeciones que podrán ser presentadas en contra de mis opiniones». Los huecos en el registro de los fósiles eran inevitables porque los fósiles sólo se forman en circunstancias excepcionales. Muchos organismos, indudablemente, llegaron y se fueron sin dejar una huella fósil de su paso y las huellas de otros pueden haber sido destruidas por catástrofes geológicas muy diversas. Pero Darwin estaba seguro, o quería creerlo así, de que algunas de las formas transitorias cruciales están enterradas y que con el tiempo y si se hacen mayores esfuerzos, acabarán por ser descubiertas. Al fin y al cabo, la Paleontología es una ciencia muy joven. «El eslabón perdido» («The mising link») era otra frase, como la de la supervivencia de los más aptos, que se apoderó del interés del público. El eslabón tenía que ser hallado o la teoría jamás llegaría a ser aceptada entre muchos científicos como entre el público. Los huesos del hombre de Neanderthal, la primera forma extinguida del hombre que llegó a ser estudiada, fueron encontrados en las cercanías de Düsseldorf, en 1856, pero los profesionales no supieron qué hacer con ellos. Incluso los partidarios de Darwin encontraron que el hombre de Neanderthal, aunque prehistórico, era demasiado humano como para considerarlo como el eslabón entre el hombre y el mono. Un eslabón perdido, que no seguiría perdido por mucho tiempo. Menos de dos años después de la publicación de la obra de Darwin, unos canteros de Baviera hicieron uno de los descubrimientos paleontológicos más fascinantes de todos los tiempos. Hallaron los restos fósiles de algo que tenía todo el aspecto de un reptil, posiblemente de un pequeño dinosaurio, que, en algunos aspectos, tenía cierto parecido con un ave, ya que poseía plumas. Darwin estableció la hipótesis de que las aves debieron de desarrollarse a partir de reptiles, y existía la prueba, al menos así parecía, en el reptil-ave conocido como el Archaeopteryx. Los partidarios de la teoría de la evolución casi no podían creer su suerte.

El descubrimiento tuvo lugar en el valle de Altmühl cerca de la ciudad alemana de Solnhofen. Desde los tiempos del Imperio romano, el valle había sido conocido por sus piedras calizas de color amarillento grisáceo, de belleza y resistencia casi marmóreas. Los canteros de Altmühl fueron descubriendo un número cada vez mayor de fósiles, algunos de ellos bastante extraños. El pterodáctilo descrito por Cuvier provenía de aquellas canteras. Otros saurios voladores habían sido igualmente preservados en la piedra, así como exquisitos especímenes de insectos, gambas, cangrejos, peces, cocodrilos y tortugas marinas. Las canteras eran una verdadera mina de fósiles para los paleontólogos. La piedra caliza de Solnhofen se había formado con los sedimentos acumulados hace 150 o 160 millones de años, a lo largo de las orillas norteñas de un mar del jurásico. El surgimiento de Europa como continente quedaba todavía a millones de años de distancia en el futuro. En aquel lugar y en aquel tiempo del jurásico, los pterosauriosplaneaban con la apariencia de enormes pelícanos pardos, a los que se parecían no sólo en su aspecto sino quizá también en la forma como se zambullían en el agua para cazar los peces. Gigantescas libélulas volaban en enjambres sobre las tierras pantanosas y las grandes charcas. A lo largo de las orillas de los ríos y los lagos, cubiertas de helechos, cicádeas y arbustos de tejo, abundaban los Compsognathus, dinosaurios del tamaño de una gallina, que corrían en persecución de pequeños mamíferos y reptiles. Y también hacían acto de presencia aquellas criaturas parecidas a las aves, los Archaeopteryx, no mayores que cornejas. F. Wemer Barthel, un geólogo crecido en la región de Solnhofen, cree que el fango fue arrastrado por tormentas hasta pequeñas bahías y lagunas y se asentó allí, detrás de una barrera de arrecifes. Las mismas tormentas, sin duda, habrían hecho caer muchas de aquellas criaturas voladoras en las lagunas, donde quedarían atrapadas en el espeso fango que en aquel tiempo formaban el lecho espeso del que se desarrollaría aquella piedra caliza de grano fino y uniforme. Al extraer la piedra de esos antiguos lechos arcillosos, los canteros trabajaban cuidadosamente para sacar las losas de piedra enteras, con tanta atención como si se tratara de piedras preciosas, por lo que tenían muchas más posibilidades de encontrar fósiles raros. John Ostrom, el paleontólogo de Yale que había visitado casi todas aquellas canteras, vio los métodos poco corrientes que allí se practicaban y que aún se siguen practicando. La mayoría de las canteras, en todas partes, están actualmente muy mecanizadas, lo que no ocurría con las canteras de Solnhofen. «Todos los trabajadores de la cantera tenían los ojos muy abiertos para descubrir cualquier trofeo fósil que, quizá, podía esconderse a los ojos de los propietarios de las canteras, y ser vendidos a un coleccionista, complementando así su escaso salario». Ostrom dijo: «Ésa es la razón por la cual en la actualidad contamos con esos especímenes increíbles del Archaeopteryx».
Fue de ese modo como los trabajadores, a finales de 1860 o principios de 1861, vieron las débiles impresiones de una pluma en uno de los bloques pétreos de Solnhofen. El descubrimiento resultó sorprendente en dos aspectos: es muy raro que texturas suaves, como las de las plumas, se conserven. Y hasta entonces nadie había encontrado el menor signo indicador de que en un período tan antiguo como el mesozoico hubiera vivido algún tipo de ave. Los aires pertenecían al pterosaurio y se suponía que las aves no aparecieron hasta después de la extinción de este saurio volador. Pero allí, en las piedras calizas de Solnhofen, del período jurásico, estaba la impresión de una pluma solitaria de unos sesenta milímetros de longitud. Que se sepa, hasta ahora las aves son las únicas criaturas que tienen plumas. Por lo tanto aquella huella fósil debió de ser dejada por el ave primitiva. La pluma fósil fue puesta a disposición de Hermann von Meyer, en Frankfurt. Era uno de los paleontólogos más respetados de Alemania. Von Meyer era el autor del libro en cinco volúmenes Fauna del mundo antiguo, que incluía sus descripciones de muchos nuevos géneros y especies de los pterosauriosdescubiertos en las canteras de Solnhofen. Era, además, autor de un precavido aditamento a la ley de Cuvier sobre la correlación de las partes. «Yo encuentro -escribió Von Meyer- que en una o en varias partes una criatura presenta correspondencias limitando con la similitud con otra criatura sin estar emparentada con ella». Los ictiosaurios, por ejemplo, pueden parecerse a los delfines en ciertos aspectos y los pterosaurios pueden parecer aves o murciélagos, pero simplemente se trata de una convergencia y no realmente de un parentesco. El reconocimiento del fenómeno de las convergencias podría haber evitado más de un disgusto a muchos paleontólogos. Eso era algo que había que tener en cuenta a la hora de examinar la pluma fósil. Von Meyer se acercó a aquel objeto con bastante escepticismo, pero finalmente llegó a la conclusión de que no se trataba de un engaño. En su informe, publicado en una revista científica alemana en agosto de 1861, declaró la autenticidad de la pluma, muy antigua y posiblemente de ave. Advirtió que «no terna necesariamente que provenir de un ave» y que podría ser de «un animal con plumas con diferencias esenciales con nuestras aves». Un mes más tarde, antes de que el mundo científico tuviera tiempo de reaccionar ante esa primera información, Von Meyer anunció otro descubrimiento mucho más sensacional. Un esqueleto sin cabeza había sido encontrado en las pizarras de Solnhofen y en él había claras impresiones de plumas. El fósil, desde luego, tenía todo el aspecto de ser uno de los eslabones perdidos preconizados por Darwin. Von Meyer le dio el nombre, neutro desde un punto de vista taxonómico, de Archaeopteryx lithographica («plumífero arcaico de la pizarra litográfica»).
El fósil llegó a manos de Andreas Wagner, de la Universidad de Munich. Aquella «criatura mestiza», observó, tenía las plumas de un ave, pero su cola era la de un reptil. Y esto le preocupaba. Wagner, un antidarwinista, se daba cuenta de lo que los evolucionistas podían hacer con ese descubrimiento y, consecuentemente, decidió golpear primero con una identificación y un nombre que desalentara a los que pensaran en hacer interpretaciones darwinianas a base del eslabón perdido. En un reportaje basado en el dibujo y en rumores, Wagner llamó al espécimen un reptil, quizá una especie de pterosaurio, y le dio el nombre deGriphosaurus problematicus, el «problemático lagarto grifo», lo cual era una declaración afirmativa, pero no una descripción taxonómica. En la mitología griega el grifo era una criatura fabulosa medio pájaro y medio león. Wagner afirmó: «La primera contemplación delGriphosaurus podrá causamos ciertamente la impresión de que tenemos ante nosotros una criatura intermedia, en la que se está operando una transformación, el paso del saurio al ave». Richard Owen, que por aquel entonces era superintendente del Museo Británico, leyó el informe de Wagner y, en 1862, envió a George Waterhouse a Pappenheim con autorización para pagar quinientas libras esterlinas por el fósil. Después de que en octubre de 1862 llegara a Londres el espécimen, Owen lo examinó con la mayor atención y detalle. Tampoco él era amigo del darwinismo. Owen creía en «la creación continua» que imitaba la evolución, pero no podía aceptar la teoría de Darwin sobre la selección natural, porque implicaba la ausencia de un designio predeterminado. Sin embargo Owen era, también, un experto anatónomo. Con una sola mirada pudo ver que aquella criatura poseía, como había dicho Wagner, «una combinación de características tan raras y sorprendentes como nadie podría imaginarse». Con la excepción de la cabeza, que faltaba, el esqueleto estaba completo y extraordinariamente bien conservado. Tenía algunas características de reptil: una larga cola huesuda; tres dedos con garras al final de sus manos delanteras, o alas; costillas y vértebras de saurio. Pero estaba claro que tenía algunas plumas, semejantes a las de las aves, que se agrupaban en las alas de la forma como ocurre en las aves modernas. Para Owen aquellas alas inclinaban la balanza. Esto, informó a la Royal Society en noviembre, era «inequívocamente» un ave antigua, pero no debía ser considerada como prueba de la existencia del eslabón perdido darwiniano. No volvió a discutir el asunto. Por su parte los evolucionistas concedieron a la criatura «importancia extrema» para documentar la teoría de Darwin. Aquel fósil era sin duda el eslabón perdido entre los reptiles y las aves. Finalmente, el darwinista Thomas Henry Huxley se sumó a la batalla y se aseguró de que el mundo conociera la importancia del Archaeopteryx.
Pese a que terna reservas sobre algunos aspectos de la teoría de Darwin, Huxley publicó un comentario favorable del libro de Darwin y se dirigió a Oxford en junio de 1860 para la primera batalla. Su presa era nada menos que el obispo de Oxford, Samuel Wilberforce, cuyas maneras untuosas le habían ganado el apodo de Soapy Sam. Wilberforce se disponía a dirigir el ataque antidarwinista en la sesión final de la Asociación Británica para el Avance de la Ciencia. Darwin esquivó el encuentro alegando que se hallaba gravemente enfermo, pero Huxley estaba allí, sentado en el estrado no lejos de Wilberforce. Entre los asistentes, unos setecientos, muchos mostraban sus simpatías por el obispo, pero no así un reducido grupo de graduados de Oxford. Según recordó Huxley, Wilberforce habló «durante una hora y media con su inimitable estilo de vaciedad y desconsideración». Posiblemente había sido entrenado por Owen, pero los relatos de la época difieren sobre cuál fue la línea de razonamiento que desarrolló en sus esfuerzos por rechazar la teoría de la evolución de Darwin. Sin embargo, todos están de acuerdo en que en su discurso, el obispo hizo una injuriosa alusión a los antepasados simiescos de Huxley, lo que llevó a éste a inclinarse hacia uno de los que se sentaban a su lado para murmurarle: «El Señor lo ha entregado en mis manos». Cuando Huxley subió al estrado desarmó por completo al obispo y se ganó el respeto, si no la completa conversión de la audiencia. En una carta a un amigo hizo este relato de su discurso: “Hablé tratando de causar el efecto de que había escuchado con gran atención el discurso del señor obispo, pero que me había sido imposible descubrir ni un nuevo argumento, ni un solo factor original en él, excepto en la cuestión surgida sobre mis predilecciones personales en cuestión de antepasados. A mí no se me hubiera ocurrido sacar a discusión un tópico así, pero estaba dispuesto a enfrentarme con el reverendo señor obispo incluso en este terreno. Si se me pusiera ante la disyuntiva de elegir, dije yo, entre tener por abuelo a un miserable mono o a un hombre altamente dotado por la naturaleza y que poseyendo grandes medios e influencia empleara esas facultades y esa influencia para introducir el ridículo en una grave discusión científica, afirmo, sin vacilación, mi preferencia por el mono“. La respuesta, de acuerdo con Huxley, desató «unas risas incontenibles» y grandes aplausos, tras los cuales la audiencia «escuchó el resto de mi disertación con la mayor atención». Más tarde Huxley incorporó al Archaeopteryx en sus ataques contra Wilberforce, Owen y otros que no se atrevían a compartir los puntos de vista de Darwin sobre el curso de la vida. Huxley estaba intrigado por las muchas características reptilianas del espécimen. Sin la impresión de la pluma, probablemente habría sido erróneamente identificado como un reptil, como había ocurrido con otros Archaeopteryx fósiles.
Unos años más tarde, en 1867, Huxley publicó un informe estableciendo la relación evolutiva entre las aves y los reptiles. Citó catorce rasgos anatómicos que se daban tanto en las aves como en los reptiles, pero no en los mamíferos, en apoyo de su caso, en defensa de clasificar a los dos juntos en una nueva categoría taxonómica a la que dio el nombre de saurópsidos. Más aún, arguyó Huxley, las criaturas más próximas, intermedias entre aves y reptiles, el eslabón de conexión que se había perdido, eran algunos tipos de dinosaurios. Huxley sentía que no había razón alguna para que los evolucionistas siguieran a la defensiva en relación a los huecos en el registro de fósiles, una vez que ya se disponía del Archaeopteryx. Expuso sus ideas sobre el tema en una conferencia popular pronunciada en febrero de 1868. Aquellos que mantienen la doctrina de la evolución, dijo el evolucionista Huxley al comenzar su discurso, «conciben que hay motivos para creer que el mundo, con todo lo que existe en él y sobre él, no comenzó a existir en la condición en que hoy lo contemplamos, ni siquiera en una forma que se aproxime en absoluto a esta condición». Continuó después con un sucinto compendio de Hutton, Lyell y Darwin, todo en uno. Los evolucionistas, dijo, «mantienen que la conformación y composición presentes de la corteza terrestre, la distribución de las tierras y las aguas y las formas infinitamente diversificadas de animales y plantas que constituyen su población presente, son meramente los términos finales de una inmensa serie de cambios que han venido ocurriendo en el transcurrir de un tiempo inconmensurable por la acción de causas más o menos idénticas a las que actúan en el momento presente». Pero ¿cómo es, preguntó Huxley en ese momento, si todos los animales «han surgido por graduales modificaciones de un mismo tronco común», que existan esos grandes «saltos», esos huecos que se dan entre peces y mamíferos o reptiles y aves, por ejemplo? «Nosotros, los que creemos en la evolución, replicamos diciendo que esos huecos fueron antaño inexistentes, que las formas que conectaban unas especies con otras existieron en épocas previas de la historia de la Tierra, pero que ahora están extinguidas». Como resultaba de todo punto natural, Huxley citó el Archaeopteryx como su primera prueba. Se mostró de acuerdo con Owen en que se trataba de un ave, pero era un ave de tipo transicional. Allí estaban las plumas y el hueso de la pechuga típico de las aves, pero también la larga cola ósea de un reptil así como otras características físicas de los reptiles que se repetían en su esqueleto.
Otro animal fósil de la misma cantera de Solnhofen, aunque contemporáneo del Archaeopteryx, fue presentado posteriormente por Huxley para robustecer su caso, no sólo en defensa del eslabón de unión existente entre los reptiles y las aves, sino de modo más específico para establecer el eslabón dinosaurio-ave. Se trataba del pequeño dinosaurio que Andreas Wagner describió en 1861 y al que dio el nombre de Compsognathus. Huxley ya se había fijado mentalmente en el Compsognathus como un eslabón perdido antes de que se uniera a los otros evolucionistas en apoyo de Archaeopteryx como tal eslabón. Si el Archaeopteryx era un ave que tenía el aspecto de un dinosaurio, el Compsognathus era un dinosaurio que, en cierto modo, parecía un ave. Tenía menos de dos tercios de un metro de longitud, el más pequeño de los dinosaurios jamás identificados y no mucho menor que el Archaeopteryx. Para Huxley resultaban del mayor interés los miembros traseros del animal, largos y delicados. El dinosaurio, por lo que parece, había sido un animal bípedo. Su articulación del pie y la del tobillo tenían un notable parecido con las de las aves. «Es imposible mirar la conformación de este extraño reptil -le contó Huxley a su audiencia- y dudar de que andaba o saltaba en una posición erecta o semierecta, más o menos como un ave, con las que debía de tener un extraordinario parecido gracias a su cuello largo, su diminuta cabeza y sus pequeños miembros anteriores». Puesto que se trataba de especies contemporáneas, Huxley reconoció que el Archaeopteryx no podía ser descendiente del Compsognathus. Pero encontrar un ave parecida a un reptil y un reptil parecido a un ave que vivieron en la misma época, aproximadamente, en el jurásico, cerraba el hueco evolutivo entre los reptiles y las clases avícolas, lo que Huxley ofreció en apoyo a su clasificación saurópsida y de la teoría de la evolución de Darwin. Del énfasis puesto por Huxley en la relación de parentesco entre los reptiles y las aves en su defensa de la teoría de Darwin surgió una nueva apreciación de los dinosaurios. Los descubrimientos anteriores fueron interpretados de nuevo. Ése fue el primero de los muchos «redescubrimientos» que haría la paleontología de los dinosaurios. Recientes hallazgos, particularmente en Norteamérica, adquirieron un nuevo significado. Los primeros fósiles hallados en Norteamérica e identificados como de dinosaurios, se excavaron en Montana y New Jersey en la década de 1850-1860. Una expedición cartográfica del Ejército de los Estados Unidos, trabajando en las tierras bañadas por el río Judith, en la actual Montana, dio con una serie de dientes, en una capa perteneciente al período cretáceo. Esos fósiles fueron examinados en 1858 por Joseph Leidy, profesor de Anatomía de la Universidad de Pennsylvania y director de la Academia de Ciencias Naturales de Filadelfia. Leidy se pronunció anunciando que aquélla era la primera prueba de que los dinosaurios también habían vivido en América durante el mesozoico. Uno de los especímenes era un herbívoro al que dio el nombre de Trachodon, lo que significa «diente rudo».

Actualmente se sabe que era uno de los dinosaurios de pico de pato que también son conocidos como Anatosaurus o «lagarto pato». Algunos hallazgos de otra criatura, en el río Judith, inspiraron a Leidy a dar a aquel carnívoro el nombre de Deinodon horridus, «el más horrible de los dientes terroríficos». En el verano del mismo año, William Parker Foulke, un miembro de la Academia de Ciencias Naturales, se enteró de la existencia de algunos huesos fósiles en una pequeña cantera en Haddonfield, en New Jersey, cerca de Filadelfia. Investigó a fondo una buena cantidad de grandes huesos, un esqueleto casi completo de una criatura de seis a ocho metros de longitud. Después de un cuidadoso examen, Leidy llegó a la conclusión que Foulke había encontrado un dinosaurio al que dio el nombre de Hadrosaurus foulkii, en honor de la ciudad y su descubridor. Se trataba de uno de los dinosaurios de pico de pato que los últimos descubrimientos demostrarían que fueron los más abundantes durante el período cretáceo en Norteamérica. Leidy dedujo de sus dientes que aquel Hadrosaurus parecía estar emparentado con el Trochodon de Montana y el Iguanodon de Inglaterra. Esto hacia pensar que los dinosaurios posiblemente habían habitado todo el mundo. Lo que más sorprendió a Leidy fue la enorme desproporción existente entre el tamaño de las extremidades anteriores y posteriores del animal. El fémur era casi dos veces más largo que el húmero del brazo superior. Si los huesos no hubieran sido hallados juntos hubiese podido creer que pertenecían a dos animales distintos. En su informe a la Academia, Leidy dijo: «La gran desproporción de tamaño existente entre las partes delanteras y las traseras del esqueleto del Hodrosaurus, me llevó a sospechar que este gran lagarto herbívoro extinguido debía de tascar y ramanear manteniéndose erecto, como el canguro, sosteniéndose sobre sus extremidades posteriores y su rabo». Huxley supo ver la importancia del descubrimiento de Leidy. No todos los dinosaurios andaban y corrían a cuatro patas. El dinosaurio rinocerino de Owen había llevado a interpretar erróneamente la realidad. Algunos de los dinosaurios eran bípedos, otro parecido generalizado entre las aves y los primates, y los dinosaurios. Seguidamente Huxley examinó de nuevo alIguanodon de Mantell y se dio cuenta de que sus cuartos traseros y su pie con tres dedos eran «sorprendentemente parecidos» a los de las aves. El Iguanodon, contrariamente a la imagen de cuadrúpedo con la que se lo representó, tenía que haber sido capaz de mantener una postura erecta y de saltar o correr sobre sus patas traseras, una observación que posteriormente sería confirmada. Una colección de huellas fósiles, algunas de las cuales tenían medio metro de longitud, llegaron a las manos de Edward Hitchcock en 1835. Se trataba de un geólogo y presidente del Amherst College. Puesto que no se halló ningún esqueleto que pudiera asociarse con las huellas, Hitchcock sólo podía imaginar cómo habían sido aquellos animales. Llegó a la conclusión de que el valle de Connecticut tuvo que ser poblado en alguna ocasión por ciertas aves extraordinarias con patas de tres dedos y de un tamaño varias veces superior al de un avestruz. «Debieron de ser los gobernantes gigantes de aquel valle», dijo Hitchcock. Nadie supo entonces que lo que se había descubierto eran huellas de dinosaurio.
Algunos dinosaurios fueron bípedos y tenían pies parecidos a los de las aves, tal como elHadrosaurus, Iguanodon y Compsognathus parecían indicar. «La importante verdad que revelan esas huellas -afirmó Huxley-, es que al comienzo de la época mesozoica existían animales bípedos que tenían pies de ave y andaban en la misma postura erecta o semierecta». Había algo que daba que pensar: los dinosaurios eran más antiguos de lo que hasta entonces se había imaginado y poseían algunos rasgos propios de las aves o semejantes a ellas aun antes de la época del Archaeopteryx. El tiempo probaría que Huxley tenía razón con respecto a las huellas. Con los nuevos descubrimientos de huesos, casi todas las huellas de Nueva Inglaterra se atribuyen en la actualidad a una variedad de dinosaurio del triásico. En el Dinosaur State Park, en Rocky Hifl, Connecticut, unas dos mil huellas bien conservadas se creen procedentes de criaturas tales como el Stegomosuchus, el Dilophosaurus y el Coelophysis, el último de los cuales, según piensan algunos científicos, ocupa un lugar destacado en la línea principal de ascendencia de las aves. Resultaba suficiente para el propósito inmediato de Huxley el haber demostrado de este modo la confusa similitud entre las huellas de las aves y las de los dinosaurios. Las aves surgieron a partir de los dinosaurios u otros reptiles arcaicos, declaró Huxley con mayor certeza que nunca, y una de las criaturas de la transición, uno de los eslabones de conexión, era el ave parecida a un reptil de las canteras calizas de Solnhofen, elArchaeopteryx. El Archaeopteryx que en la actualidad se encuentra en el museo de Londres no tenía cabeza y algunos de los antidarwinistas se aferraron a ello con todas sus fuerzas. Si elArchaeopteryx hubiese tenido un pico sin dientes, como todos deseaban creer, entonces hubiera sido seguramente un ave, aunque antigua. Incluso Huxley dudaba de que el ave primitiva tuviera dientes, aunque otros que habían examinado el fósil creyeron ver los restos de un maxilar superior con cuatro dientes. Si hubiera sido una boca de reptil con dientes, entonces Huxley y sus aliados se hubieran sentido reforzados en su hipótesis de que se trataba de un eslabón evolutivo entre reptiles y aves. Aquel pájaro sería de mucho más valor si hubieran podido encontrar otro con cabeza. El siguiente descubrimiento se produjo en 1877, también en la región de Solnhofen, y aquel esqueleto estaba aún más completo y era más espléndido que el primero.
Alan Feduccia, un zoólogo norteamericano, escribió en 1980 para decir que este fósil deArchaeopteryx «posiblemente será el más importante espécimen de la historia natural, tal vez comparable con el valor de la piedra Rosetta». Su cabeza estaba intacta, una cabeza de Huxley. El cráneo era de ave con una boca que se desarrollaba hacia adelante adquiriendo forma de pico, pero en las mandíbulas había dientes afilados de reptil. En contraste con el espécimen de Londres este esqueleto estaba articulado en una pose natural con las alas extendidas. Los brazos y manos de tipo dinosaurio, estaban adornadas con una gran cantidad de plumas casi idénticas en detalle a las de las aves modernas y había más plumas a ambos lados de su larga cola de reptil. En esta ocasión, el Archaeopteryx se parecía aún más al eslabón perdido. El fósil fue clasificado como si fuera un género especial, Archaeomis siemensi. Sin embargo, estudios posteriores demostraron que el segundo espécimen pertenecía probablemente al mismo género, si no a la misma especie que el primer Archaeopteryx. Entre los paleontólogos el segundo Archaeopteryx, en la actualidad propiedad del Museo de la Universidad Humboldt, en Berlín, es mencionado generalmente como el espécimen de Berlín. Desde entonces han sido identificados otros esqueletos de Archaeopteryx. John Ostrom, mientras se hallaba en Europa para estudiar los especímenes de Londres, Berlín y Maxberg, también visitó el Museo Teyler, en Harlem, Países Bajos. Allí encontró un espécimen que había sido clasificado como unpterosaurio, pero que no lo era en absoluto. De inmediato se dio cuenta de que las piernas y los pies eran demasiado largos para un pterosaurio de aquella era. Los tres dedos de las garras eran de distinta longitud mientras que una de las características del pterosaurio es que son igualmente largos. Ostrom se sintió picado en su curiosidad y consiguió permiso para sacar el fósil de la vitrina en que se exhibía para poder inspeccionarlo más de cerca y detalladamente. Cuando colocó la losa de piedra caliza bajo una luz fuerte pudo ver leves impresiones de plumas, algo que nadie había descubierto previamente cuando se realizó el hallazgo, porque no existía razón que los pudiera llevar a sospechar la existencia de algo tan extraño. Los pterosaurios eran reptiles voladores, o planeadores, con alas de membranas de piel, no de plumas. Ostrom le preguntó al director del museo de Teyler si podía prestarle el fósil para llevárselo a su casa, en Yale, y poder estudiarlo con detalle. Explicó su sospecha de que en realidad el pterosauriopodía ser un Archaeopteryx, algo que cualquier director de museo se sentiría satisfecho de oír. Hermann von Meyer, que lo investigó, observó que la «membrana de las alas» tenía una extraña apariencia, distinta a la de otros pterosaurios que él había visto. Pero se limitó a informar que se trataba de una especie distinta de pterosaurio.

Ostrom, de vuelta en Yale en el otoño de 1970, confirmó su primera impresión de que se trataba de un nuevo fósil de Archaeopteryx y anunció el redescubrimiento, para decirlo así, de un nuevo ejemplar. Aunque el espécimen de Teyler «está en estado fragmentario y es mucho menos espectacular» que los ejemplares de Londres y de Berlín, dijo Ostrom, tiene garras bien conservadas que ofrecen «nuevas evidencias importantes que no están conservadas en ninguno de los otros especímenes». Una especie de vaina o cubierta córnea, en cierto modo semejante a una uña humana, rodeaba las garras. Así, pues, ahora se dispone de cinco especímenes de esta rara ave primitiva: los ejemplares de Londres, Berlín, Maxberg, Teyler y Eichstatt, así como la pluma solitaria que se conserva en el museo de Munich. Othniel Charles Marsh declaró en 1877 que Huxley, realmente, había establecido el puente entre reptiles y aves. El Compsognathus y el Archaeopteryx, dijo, sirven como «escalones de piedra por los cuales los evolucionistas de hoy día conducen al dudoso hermano para cruzar los vacíos restos del golfo, que antaño pareció insalvable». Durante varios años, la hipótesis de que las aves eran descendientes directas de los reptiles gozó de amplio apoyo. El Archaeopteryx es, todavía, el mejor ejemplo conocido del tipo de criaturas predecidas por la teoría de Darwin: una especie que representa un paso intermedio entre dos grandes clases de organismos. El descubrimiento del Archaeopteryx, que tuvo lugar poco después de que Darwin publicara su libro The Origin of Species, elevó el estudio de los dinosaurios a un plano más amplio y más elevado. Pasaron a ser considerados algo más que monstruosas curiosidades. Era más que posible que los dinosaurios fueran figuras claves en algunas evoluciones trascendentales en el curso de la vida en la Tierra. Desde el Oeste americano llegaron, en 1877, tres mensajes sobre descubrimientos de huesos poco corrientes que habrían de convertirse en objeto de controversia y en la más enconada de las rivalidades en la historia de la paleontología. Pusieron en movimiento expediciones exploradoras que descubrieron algunos de los más ricos cementerios de dinosaurios del mundo y llevarían al descubrimiento de unas criaturas del pasado tan populares como el Brontosaurus y el Diplodocus, el Stegosaurus y el Triceratop. Los nuevos hallazgos revelaron, por encima de toda duda, que los dinosaurios vivieron en gran número en el mesozoico y que existieron en una variedad insospechada de formas y tamaños.En noviembre de 1877 se recuperaron unas treinta toneladas de restos de dinosaurios. Había huesos muy raros y esqueletos parciales deCamarasaurus, Laosaurus, Dipiodocus, Barosaurus y Stegosaurus. El reparto de papeles entre los dinosaurios se iba haciendo cada vez más definido. Los fósiles eran abundantes y mezclados.
Charles F. Stemberg era uno de los cazadores de dinosaurios más destacados, entre los que trabajaron en las excavaciones para Cope o Marsh, y que continuaron realizando el mismo trabajo después de que los dos eternos rivales abandonaron. Al igual que la mayor parte de aquellos buscadores de fósiles, Stemberg era un hombre del Oeste, amante de la vida en el campo, al aire libre. En respuesta a la llamada de los fósiles, se convirtió en una especie de cowboy o de buscador de oro, salvo que en vez de oro buscaba fósiles. Stemberg descubrió una momia de dinosaurio. Se trataba de uno de los dinosaurios de pico de pato conocidos por el nombre de trachodontes. Cuando el animal murió, hacía unos sesenta y cinco millones de años, el cuerpo se momificó bajo condiciones de extrema sequedad, en las que las humedades se evaporaron antes de que las bacterias hubieran llegado a producir la descomposición. De algún modo logró escapar a los carroñeros y rápidamente se cubrió con arenisca y se fosilizó posteriormente. Y se quedó allí, piel, tendones y trozos de carne petrificados hasta que los hombres de Stemberg dieron con él. Las praderas del oeste del Mississippi están llenas de los pequeños montículos de los habitáculos de la hormiga carpintera roja. Excavando el interior de sus hogares subterráneos las hormigas encontraban trozos de huesos y de piedras que sacaban y dejaban fuera. Acumulaban aquellos fragmentos encima de sus montículos. También se dice que esas hormigas se salen de su camino para buscar objetos pequeños y duros que llevan de vuelta a sus montículos para fortalecerlos. Los científicos no están seguros de por qué motivo las hormigas hacen esto. Es posible que ese material tosco y duro proteja los hormigueros contra la lluvia y la erosión del viento. También es factible que las piedrecitas y los pequeños huesos sirvan para el almacenaje del calor solar, que ayude a la incubación de los huevos de las hormigas y a las larvas que ocupan el interior de los montículos. Pero lo único que verdaderamente le importaba a Hatcher era que muchos de los hormigueros contenían una buena cantidad de pequeños huesos fósiles y de dientecitos. Las hormigas habían realizado la mayor parte de su trabajo. Hatcher utilizaba una pequeña criba de las que sirven para cerner la harina para separar los huesos de la tierra polvorienta. Los actuales paleontólogos se alegran cuando descubren la presencia de hormigueros y los registran hasta el punto de haber encontrado en ellos miles de pequeños huesos de mamíferos que han revelado muchas cosas de esos diminutos contemporáneos de los últimos dinosaurios. Cuando Bamum Brown llegó en 1897 al Museo Americano, éste no poseía ni un solo fósil de dinosaurio. Cuando murió Brown en 1963 -sólo le faltaba una semana para celebrar su nonagésimo aniversario- el museo tenía una de las más extensas e importantes colecciones del mundo, lo que se debía, en gran parte, a las numerosas exploraciones a gran escala realizadas a lo largo de sesenta años. Su descubrimiento más impresionante fue el de los primeros esqueletos de Tyrannosaurus rex, el mayor carnívoro que cazó sobre la tierra firme. Brown realizó su hallazgo más sensacional en el peñasco de dura piedra caliza amarillenta que cruzaba un arroyo.
Brown encontró los primeros huesos de lo que, según se vio después, era el esqueleto completo de un Tyrannosaurus rex. Las partes mayores del esqueleto, como la pelvis, incrustada en la piedra, que pesaba en su conjunto unas dos toneladas, significó una auténtica prueba para demostrar la capacidad estratégica de Brown. Dado que resultaba demasiado pesada para poder ser llevada por el carro, construyó una especie de trineo con tablones deslizantes y alquiló un tiro de cuatro caballos para arrastrarla. Costó todo un día de trabajo trasladar la pelvis desde el lugar en que fue hallada, en el valle del arroyo del Infierno, hasta la carretera. Allí comenzaba el transporte durante doscientos kilómetros de distancia hasta la estación de Miles City. Cinco años más tarde, en 1908, Brown descubrió su segundoTyrannosaurus. De regreso al museo de Nueva York, los huesos de los dos especímenes de tiranosaurio fueron ensamblados con el máximo cuidado para que el mundo pudiera contemplar el terrorífico aspecto que debieron de tener los más poderosos carnívoros terrestres que jamás vivieron. Esas criaturas, que existieron en los últimos millones de años del cretáceo, tenían cabezas enormes, de más de un metro de longitud, desde el frente hasta la parte de atrás. Las mandíbulas apenas si eran algo más cortas y estaban llenas de dientes y colmillos de sable de quince centímetros de longitud y de dos centímetros y medio de anchura. Esos animales se erguían sobre sus dos patas traseras que terminaban en pies semejantes a los de las aves, con tres dedos dirigidos hacia adelante, armados con fuertes espolones, las armas de las que se valían para apresar sus presas. Entre los más valiosos especímenes encontrados por Brown se cuentan un esqueleto casi completo de un dinosaurio con cuernos, el Monoclonius, y otro del dinosaurio de pico de pato y con cresta llamado Corythosaurus. Y lo que era más interesante para los paleontólogos, Brown estuvo en condiciones de extraer fósiles de distintos estratos del cretáceo y con ello reveló nuevas pruebas del desarrollo y la evolución del dinosaurio durante los últimos millones de años de su existencia. Un año después de los sorprendentes descubrimientos de 1877 en el Oeste americano, los trabajadores que estaban abriendo una nueva galería de más de trescientos metros en una mina de carbón belga, encontraron un rico yacimiento de huesos. Habían hecho un túnel hasta llegar a una capa cretácea que profundizaba atravesando los depósitos carboníferos formados por los bosques pantanosos del más antiguo período carbonífero. Una profunda barranca debió de abrirse a través del carbón, unos cien millones de años antes, y en ella cayeron muchos dinosaurios indefensos cuyos cuerpos serían pronto cubiertos por las tierras pantanosas y las aguas de las inundaciones. El barro se petrificó y los huesos se fosilizaron apresados en él. Los fósiles se quedaron allí hasta que los seres humanos de la era industrial necesitaron carbón. Los mineros, sin querer, destruyeron un esqueleto antes de darse cuenta de que habían dado con un cementerio prehistórico.

Seguidamente y durante los tres años siguientes los paleontólogos del Museo Real de Historia Natural en Bruselas trabajaron en la sombría oscuridad de la galería minera en Bemissart, cerca de la frontera francesa. Extrajeron de allí más de treinta esqueletos, virtualmente completos, de Iguanodon. Nunca antes se habían encontrado tantos especímenes de un solo género de dinosaurios en un mismo lugar, una circunstancia que ofrecía a los científicos la rara oportunidad de estudiar las variaciones de una especie y de recrear las vidas y el hábitat de los dinosaurios. Louis Dollo fue un paleontólogo que hizo una carrera ilustre gracias a esos Iguanodon de Bemissart. En el término de un año, Dollo ensambló y montó su primer esqueleto y comenzó a publicar sucintas interpretaciones sobre cuál debió de haber sido el aspecto y la vida de aquellos animales. Trabajando con esqueletos completos, Dollo pudo informar con certeza que elIguanodon se erguía y se movía sobre sus patas traseras y con las manos delanteras libres para coger hojas y ramas de los árboles, así como otras plantas comestibles. El Iguanodon no era un cuadrúpedo, sino un bípedo, como había postulado Huxley. De adulto el Iguanodon era un animal fuerte, grande, que posiblemente llegaría a pesar siete toneladas, con unos diez metros de longitud y cinco de altura. Dollo mostró, además, que la protuberancia ósea, que Mantell había tomado por un cuerno semejante al que va sobre el hocico del rinoceronte, era una gran garra, una especie de espolón, que iba en la punta del dedo pulgar del Iguanodon.Posiblemente era una de las armas defensivas del colosal herbívoro. Al examinar la cola delIguanodon, Dollo descubrió una ordenación de tendones osificados que cruzaban las vértebras y supuso que su misión consistía en dar rigidez a la cola que debía de tener consecuentemente una fuerza que anteriormente no se había supuesto. Una cola rígida podría haber sido el arma de combate más efectiva del dinosaurio. Bastante tiempo después los científicos llegaron a la conclusión de que aquella cola debía de servir también para que el animal pudiera mantener el equilibrio cuando corría saltando sobre sus dos patas traseras.En sus estudios del dinosaurio, Dollo avanzaba en un terreno al que dio el nombre de paleontología etológica. Miraba más allá de los huesos para ver cómo habían vivido los dinosaurios. Naturalmente que la anatomía de aquellos animales era una de las claves más importantes, puesto que de ella se podía deducir su postura, sus posibles hábitos alimenticios y su defensa contra los depredadores. Además, Dollo quiso recrear el medio ambiente en el que había vivido el dinosaurio. Informó sobre los fósiles de los animales, insectos y plantas que compartieron el mundo del Iguanodon. Estudió los sedimentos para deducir cuál debió de ser el clima y el paisaje de aquel período de la antigüedad. Dollo fue pionero en muchos de los métodos que se desarrollarían con creciente sofisticación en el siglo XX y que nos han dado una increíble visión del pasado.
En lo que se refiere a los Iguanodon de Bemissart, de acuerdo con la recreación documentada de Dollo, debieron de haber pastado en un extenso delta fluvial que, en aquella era, cubría Bélgica, el norte de Francia y el sur de Inglaterra, entonces unidas, hace unos cien millones de años, en el cretáceo. El delta se abría en un amplio mar que se extendía a través del sur de Europa hasta Asia, el mar Thetis. El clima era tropical. Las coniferas crecían sobre el suelo cubierto de helechos. Entre los cocodrilos, tortugas, ranas y grandes insectos voladores, losiguanodon vivían en gran número y pastaban, siempre alertas, vigilando cualquier señal que pudiera anunciar la presencia de los dinosaurios carnívoros depredadores. El Iguanodonposiblemente se defendía del ataque de los carnívoros con sus garras y con su cola flagelante o, quizá, huían a refugiarse en las aguas. Algunos de ellos, por una razón ignorada, quedaron atrapados en la barranca profunda que les sirvió de tumba. Louis Dollo llegó a ser profesor en la Universidad de Bruselas y amplió sus estudios a otros dinosaurios, entre los que se incluían elTriceratops y el Stegosaurus. Para el momento de su muerte, en 1931, ya se habían desenterrado muchos otros dinosaurios en Europa, desde Escocia hasta Transilvania, desde los Países Bajos a España y Portugal. Pero ninguno de los descubrimientos hechos en Europa, antes o después de su muerte, admite comparación con los esqueletos de Iguanodonencontrados en la mina de carbón de Bemissart. En su búsqueda de dinosaurios, como en otras muchas cuestiones, los europeos no podían limitarse a su propio continente. Los colonos británicos en África del Sur enviaban en ocasiones a Londres huesos misteriosos para que fueran estudiados. El primero de ellos, identificado como perteneciente a un dinosaurio, fue un espécimen de un prosaurópodo del triásico, y fue descrito por Owen, en 1854, que le dio el nombre de Massospondylus. En un principio se pensó que esos animales, cuya longitud más común era de unos cuatro metros, podrían ser los antepasados directos de los grandessaurópodos del jurásico, cuadrúpedos gigantescos como los Brontosaurus. Alan Charing afirma que ahora son muchos los especialistas que creen que los prosaurópodos eran una rama colateral del árbol de la familia de los saurópodos que se extinguió a finales del triásico. «Los antepasados del triásico de las mayores bestias que jamás corrieron por la Tierra todavía tienen que ser descubiertos», dijo Charing. Otras reliquias que llegaron a Londres procedentes de África del Sur, en 1866, frieron identificadas por Huxley como una nueva especie deprosaurópodos del triásico, el Euskelosaurus. Muy poco más se oyó hablar de los dinosaurios africanos hasta que Robert Broom comenzó a informar de sus exploraciones a principios de 1900. Broom, médico escocés, que estudiaba los orígenes de los mamíferos, había trabajado durante años en Australia investigando la anatomía de algunos mamíferos primitivos como el ovíparo monotremes y los marsupiales antes de que se trasladaran a África del Sur.
Allí, su interés recayó de modo principal en los restos fósiles de algunas raras criaturas conocidas como reptiles de aspecto de mamíferos. De algunos de esos animales, que vivieron en el período pérmico y triásico, se desarrollaron los pequeños mamíferos que lucharon por sobrevivir durante la era de los reptiles. Tal vez fueros los ancestros del hombre. Años antes de que se estableciera que África fue la cuna de la especie humana, Broom encontró, en el Gran Karoo, una cuenca árida situada en el extremo sur del continente, numerosas pruebas de la existencia de los primeros antepasados del primer mamífero. Mientras cazaba con trampas por el Karoo, halló algunos huesos de dinosaurio sobre los cuales envió su primer informe en 1904. Broom describió los huesos de un Brontosaurus, el Algoasaurus, que vivió a principios del período cretáceo. Esto dio ánimos a los paleontólogos para empezar la caza de dinosaurios en África. Muy pronto encontraron todos los huesos que habían confiado en hallar, en un lugar llamado Tendaguru. Los lechos de fósiles de la región demostraron ser aún más ricos que todos los encontrados en el hemisferio sur. En 1907, un ingeniero de minas alemán, W. B. Sattler, que trabajaba a cien kilómetros en el interior del país, a partir de Lindi, un puerto marítimo del África Oriental Alemana, que en la actualidad se llama Tanzania. Cerca de Tendaguru, una colina que se alza frente a una pradera cubierta de hierba y de árboles pequeños, Sattler encontró varias piezas de huesos largos que yacían sobre el suelo y que reconoció como fósiles. Eberhard Fraas, una autoridad en reptiles fósiles, que se hallaba por casualidad en la colonia, de visita, fue llevado al lugar del hallazgo para que identificara aquellos huesos. Por todas partes que dirigió su vista entre la alta hierba, pudo ver más huesos, que habían sido puestos al descubierto por la erosión. Después de recoger algunas muestras regresó a su casa, en Stuttgart, e hizo correr la voz de la existencia de un fabuloso yacimiento de huesos de dinosaurio que simplemente esperaba la llegada de alguien con pico y pala. Se pudo reunir el suficiente dinero para financiar una expedición a Tendaguru. Bajo la dirección de Wemer Janensch, el encargado de la sección de reptiles en el Museo de Berlín, paleontólogos alemanes y cientos de personas de la localidad, trabajaron en aquel lugar durante cuatro años a partir de 1909. Fue una empresa monumental, mucho más que cualquiera de las expediciones en busca de dinosaurios en el Oeste americano. En primer lugar allí no había ferrocarril y el campamento estaba a dos días de distancia, en buque fluvial, de Dar-es-Salaam, al sur de Lindi, un campamento caluroso muy poco recomendable para los visitantes europeos. Desde Lindi era precisa una marcha de cuatro días, en el calor tropical del interior, para llegar a Tendaguru.

El más precioso de los trofeos conseguidos en esta expedición fue un gigante del jurásico, elBrachiosaurus -o lagarto con brazos-. Cuando, de regreso en Berlín, Janensch ensambló los huesos de aquella criatura el resultado fue realmente impresionante. Aparentemente se trataba del mayor de todos los cuadrúpedos, saurópodos herbívoros, el formidable dinosaurio que hoy nos resulta tan familiar. A diferencia de la mayor parte de otros dinosaurios, el Brachiosaurustenía las patas delanteras mucho mayores que las traseras. Su espalda descendía en pendiente hacia atrás y surgía de un cuello larguísimo al final del cual había una cabeza relativamente pequeña. Esta cabeza quedaba a 12,6 metros del suelo, es decir, a suficiente altura para que elBrachiosaurus pudiera mirar por encima de una casa de cuatro pisos. Janensch calculó que el animal vivo debía de pesar unas dieciocho toneladas, hasta veinte veces el peso del mayor de los elefantes africanos. En la actualidad el esqueleto del Brachiosaurus se exhibe en el Museo de Historia Natural de la Universidad Humboldt en Berlín. Es, sin duda, el resultado más espectacular de la expedición a Tendaguru y el mayor testimonio de la larga carrera científica de Wemer Janensch. Hasta fecha muy reciente se había considerado al Brachiosaurus como el mayor de los animales que existieron en la Tierra en todos los tiempos. Pero otro dinosaurio, quizá un pariente distante del Brachiosaurus, es posible que lo sobrepasara en peso y altura. Se trata del llamado Ultrasaurus, que Jim Jensen encontró en Colorado en 1979. Partiendo del tamaño del omoplato, la única pieza grande del esqueleto que ha sido desenterrada, Jensen calcula que este dinosaurio tenía, por lo menos, un piso más de altura que el Brachiosaurus de Tendaguru. En 1930, en su libro The Dinosaurs in East Africa, Parkinson atribuye la concentración de dinosaurios en Tendaguru a las probables condiciones que reinaron allí en el último período del jurásico. Los dinosaurios vivían en la amplia desembocadura de un río que vertía sus aguas en una laguna separada del océano por un gran banco de arena. «No cabe duda de que muchos animales serían apresados en aquella laguna y en ella hallaron su tumba», escribió Parkinson, trazando una imagen no muy agradable. «Las carcasas en estado de podredumbre flotaban gracias a los gases producto de la descomposición y se iban desmembrando lentamente -continuaba-. Las arenas en las cuales se encuentran los huesos son invariablemente de grano fino y para que tales pesos pudieran ser transportados por el agua, que necesariamente tenía que transcurrir a poca velocidad, cabe suponer que el conjunto era una masa fangosa, un verdadero barrizal». En 1929, los británicos extendieron las exploraciones un poco más hacia el norte, hasta un lugar llamado Kindope. Allí, entre otros varios fósiles, los británicos encontraron un dinosaurio que se parecía al Stegosaurus escamoso de Norteamérica, aunque un poco más pequeño y con una altura de unos cinco metros. Se le dio el nombre de Kentrosaurus.
Los cazadores de fósiles habían establecido la distribución global de los dinosaurios. El primer lugar donde se excavaron sus huesos fue en Inglaterra, después en el resto de Europa, de modo especial en la cuenca minera de Bemissart, en Bélgica. Después fueron encontrados en gran abundancia en Norteamérica, de modo muy especial en los estados y las regiones del oeste. En 1882 se identificaron los primeros huesos de dinosaurios en América del Sur, exactamente en Argentina. En 1924, algunos especímenes de dinosaurios fueron sacados a la luz en Australia. Y un verdadero tesoro de restos fósiles que esperaban en Asia, inspiraron una nueva aventura en el campo de la caza del dinosaurio. A primeras horas de la mañana del 21 de abril de 1922, un grupo de hombres dejó la ciudad fronteriza de Kalgan, al norte de Pekín, y se encaminaron hacia la puerta de la Gran Muralla detrás de la cual está Mongolia. Innumerables caravanas habían hecho aquella misma ruta con anterioridad, viajeros de todo tipo y aspecto, pero sin duda ésta era la más extraña de todas. En el grupo iban varios científicos norteamericanos, amantes de la aventura, naturalistas, paleontólogos y geólogos, con toda una cohorte de mecánicos, cocineros y ayudantes. Días antes los había precedido una caravana de setenta y cinco camellos que transportaban bidones de gasolina y cajas con otros suministros, que debían ser depositados en los lugares designados de antemano a lo largo de la ruta por la cual se intentaba cruzar el Gobi, ese gran desierto que se extiende al este y al oeste, durante 3 000 kilómetros, en el centro de Mongolia. Aquellos cuarenta hombres, cinco vehículos a motor y setenta y cinco camellos, formaban la expedición del Asia Central, organizada y dirigida por Roy Chapman Andrews. Éste describió la empresa como «la mayor expedición científica terrestre que jamás salió de Estados Unidos». Nada semejante se había visto nunca en Asia, ni figuraba en los anales de la caza del dinosaurio. El principal objetivo de esta expedición no tenía nada que ver con el dinosaurio, sino que se trataba, nada menos, que de la búsqueda del origen del hombre. Tan ambicioso proyecto hubiera despertado sospechas entre los incrédulos burócratas chinos y mongoles. No podían creer que hombres en su sano juicio se arriesgaran a cruzar el desierto de Gobi simplemente para buscar viejos huesos. Aquella historia tenía que ser la cobertura, el pretexto, tras el cual se ocultaba otra cosa. Supusieron que su objetivo tendría que ser algo más valioso, como el oro o el petróleo, o quizá tenían la intención de apoderarse de todo el país y aquellos hombres que se llamaban a sí mismos científicos no eran otra cosa que exploradores en busca de tesoros o algo aún peor, agentes del imperialismo norteamericano. Esto era algo que podían entender, aunque no tolerar, pues esas razones eran las que durante muchos años venían atrayendo a los extranjeros a China y ahora a Mongolia. Pese a todo, debido a las cartas de presentación de altos funcionarios norteamericanos y a la intervención de gente influyente en Pekín, Andrews obtuvo el permiso necesario y se puso en acción tan pronto la nieve empezó a fundirse y el calor de la primavera volvió a Gobi.
En el año que Andrews había pasado en Pekín ocupado con los preparativos de la expedición, había tenido que oír los consejos de muchos que le prevenían en contra de su proyecto de partir de Kalgan en dirección a Urga, en la actualidad Ulan Bator, la capital de Mongolia. Recordó que muchos le habían dicho que debían de estar casi locos. Gobi era una tierra desértica, estéril, de arena gruesa, salvia camellera y arbustos espinosos. En el verano, durante el día, el calor se hacía realmente insoportable. Pero eso, con ser muy desagradable, no era nada en comparación con los vientos gélidos con los que habrían de enfrentarse si se veían obligados a pasar allí el invierno. También era posible que el elemento humano fuese igualmente inhospitalario. Los bandidos amenazaban continuamente la ruta de las caravanas y las luchas políticas azotaban el país. Mientras Andrews invernaba en Pekín, los ejércitos de Mongolia habían logrado expulsar a los chinos fuera de sus tierras. En muchas ocasiones su expedición pasaría por lugares que evidenciaban la existencia de grandes batallas que no figuraban en los archivos y de las que el mundo exterior no supo nada. Apenas los mongoles vieron su país libre de chinos, cuando contemplaron cómo los rusos se infiltraban por el norte y el oeste. Las tropas bolcheviques habían entrado en Mongolia en persecución de unidades de rusos blancos que no querían rendirse. Pero, después de haber acabado con ellos, los bolcheviques se quedaban allí para aprovecharse del caos político que había seguido a las guerras chinas. En la década de los años veinte Mongolia, gradualmente, fue cayendo bajo el control ruso; precisamente en el período en que Andrews estaba allí buscando el origen del hombre y cualquier otro tipo de huesos prehistóricos que encontrara en su camino. La historia fósil de partes alejadas de Asia, incluso Mongolia, era completamente desconocida. El desierto de Gobi había sido cruzado en ambas direcciones y en múltiples ocasiones por varios exploradores y naturalistas en los años finales del siglo XIX, pero como no tuvieron demasiado tiempo para una exploración a fondo, se habían dado por satisfechos con una inspección superficial. Si encontraron algún fósil no informaron de ello, salvo con una excepción: en 1892 el geólogo ruso Vladimir A. Obruchev encontró un diente de rinoceronte fósil cuando su caravana hacía la ruta de Kalgan a Urga. En 1900 Henry Fairfield Osbom había profetizado que en el Asia Central los paleontólogos encontrarían la cuna original del hombre primitivo. La idea había ganado cierta respetabilidad científica unos dos lustros antes, cuando William Diller Matthew, director de la sección de paleontología de los vertebrados en el Museo Americano de Historia Natural, postuló en un estudio sobre el clima y la evolución que el Asia Central podría muy bien ser el Jardín del Edén de la mayor parte de los mamíferos, incluso quizá del propio ser humano. Se trataba de una hipótesis que merecía ser estudiada sobre el terreno. Ésta era la razón por la que Andrews entró en escena, un hombre en busca de aventuras y determinado

Andrews quiso ir a la exploración del Gobi, debido a la influencia de Osbom y Matthew. La idea de buscar los orígenes del hombre en una tierra extraña y remota disparó su imaginación, pero no cruzó por su mente la idea de buscar dinosaurios. En un lugar tan árido e inhóspito como el desierto de Gobi no existía posibilidad alguna de vivir sobre el terreno. De vez en cuando, la expedición podría comprar un cordero, cazar un antílope o aprovisionarse en Urga, pero no se podía confiar en ello. Todas las necesidades debían ser tenidas en cuenta de antemano y había que llevar consigo todas las provisiones necesarias. La expedición de Andrews tuvo unos buenos momentos después de pasar por la puerta de la Gran Muralla y en cuatro días la caravana motorizada recorrió 425 kilómetros desde Kalgan, es decir, la mitad de la distancia entre Kalgan y Urga. El equipo alcanzó la laguna de Iren Dabasu, un terreno más estéril que el que habían dejado atrás, si es que eso era posible. Montones de arena, de forma cónica y de color rosado, se alzaban sobre el suelo como gigantescos hormigueros. En tiempos más tranquilos y pacíficos, obreros chinos reclutados en Kalgan recogían la sal del lago y la enviaban al sur en caravanas de carros tirados por bueyes. Pero aquéllos no eran tiempos de paz y cuando Andrews llegó allí se encontró con una escena de la máxima desolación. Andrews decidió establecer un campamento en la base de unos peñascos de un color gris blancuzco. Mientras supervisaban la instalación de las tiendas, Granger, Berkey y Morris recorrieron los alrededores en coche para inspeccionar la zona. A la caída del sol, los dos coches de los científicos dieron la vuelta en tomo a un gran peñasco pardo y regresaron al campamento. Andrews se quedó mirando a Granger y vio que sus ojos brillaban de emoción. Berkey estaba «extrañamente silencioso». Granger metió la mano en sus bolsillos y sacó un puñado de fragmentos óseos. Berkey y Morris hicieron lo mismo. A la mañana siguiente, antes del desayuno, los científicos dejaron el campamento para buscar nuevos fósiles. Berkey encontró un fragmento del fémur de un animal que no supo identificar. A los ojos de Granger era de aspecto reptil. Granger subió a la parte más alta del macizo rocoso y cayó de rodillas. Comenzó a barrer la arena que cubría algo empotrado en el suelo. Era la tibia de un gran reptil. Lo que habían encontrado eran los huesos de un dinosaurio, el primero hallado en Mongolia y uno de los primeros localizados en cualquier otro lugar del Asia oriental. Un Andrews pletórico de júbilo pudo olvidarse de los escépticos que allí, en Pekín, se habían reído de él. Allí estaban los fósiles de Mongolia y había sido él quien dio con ellos. Los fósiles eran mucho más antiguos que el ser humano o que cualquier otra cosa que hubiese esperado encontrar. Reagrupada de nuevo, la expedición regresó al desierto de Gobi, en esta ocasión por el sudoeste de Urga, y se dirigió a los lugares donde debería realizar sus descubrimientos más importantes. Al día siguiente a su llegada al campamento del Onagro, Shackelford, el fotógrafo, encontró un hueso muy bien conservado perteneciente al pie de una especie de antiguo rinoceronte.
Andrews buscó y excavó por los alrededores y descubrió algunos dientes medio rotos. A Granger le costó cuatro días de meticuloso trabajo extraer los dientes del suelo y el cráneo al cual pertenecían. El hueso del pie encontrado por Shackelford y el cráneo resultaron ser los hallazgos más interesantes de la expedición. De regreso en 1924, Osbom determinó que habían pertenecido al rinoceronte gigante extinto llamado Baluchitherium, que anteriormente sólo había sido descubierto en Baluchistán. Se trataba del mayor de los mamíferos terrestres que jamás existió en nuestro planeta, por lo que hasta ahora se sabe. Regresando hacia el este para comenzar el viaje de vuelta a China, la expedición siguió una antigua ruta de caravanas que una tarde los llevó al borde de un amplio lago, rodeado por paredes de piedra arcillosa de color rojizo. El paisaje resultaba impresionante. Los hombres, asombrados y maravillados, lo bautizaron con el nombre de Flaming Cliffs («Acantilados Flamígeros») de Shabarakh Usu, actualmente Bain-Dzak. Andrews se quedó extasiado: «Desde nuestras tiendas podíamos ver bajo nosotros un extenso estanque rosado, tachonado con salientes rocosos como extraños animales esculpidos en las rocas. Una de aquellas peñas fue bautizada con el nombre de “dinosaurio”, porque parecía un gigantesco Brontosaurus, sentado sobre sus patas traseras. Otras parecían ser castillos medievales con almenas y torretas, como de ladrillo rojo a la luz del atardecer, puertas colosales, muros y terraplenes. Las cavernas se hundían profundamente en las rocas, y laberintos de barrancos y gargantas estaban llenos de fósiles que los convertían en un paraíso para los paleontólogos». Shackelford se dirigió directamente, sin vacilar, hacia un pequeño pináculo de rocas, en la cima de las cuales descansaba un hueso fósil blanco. Cuando Granger examinó aquel fósil llegó a la conclusión de que se trataba de un trozo del cráneo de un reptil. Fue recogido con sumo cuidado. Después, todos los hombres descendieron a la laguna y se dispersaron, removiendo rocas y arena, para regresar con todos los fósiles que podían llevar consigo. Entre otras cosas Granger aportó un trozo de cáscara de huevo. Los buscadores apenas si le concedieron importancia, limitándose a creer que se trataba de la cáscara de un huevo de un ave fósil. Al año siguiente sabrían la verdad. De mala gana Andrews abandonó Flaming Cliffs, prometiendo regresar. «Difícilmente podíamos suponer en aquel entonces, que más adelante llegaríamos a considerarlo el más importante depósito en Asia, si no en el mundo entero», escribió. Pero a medida que pasaba el tiempo el viento soplaba cada vez más frió y la expedición tuvo que regresar a China en octubre. La fortuna les había sido propicia: habían encontrado sus primeros fósiles al principio del viaje, habían sabido negociar con éxito sus permisos y visados con las autoridades administrativas de Urga, y habían quedado impresionados por el esplendor de la promesa paleontológica en Flaming Cliffs. No habían tenido problema alguno, salvo algunas pequeñas dificultades con los vehículos a motor y, de un modo u otro, habían eludido todo encuentro con bandidos o guerrilleros.
En ese mismo invierno, Osbom cablegrafió: «Ha hecho usted un descubrimiento muy importante. El reptil es el antecesor del Triceratops tanto tiempo buscado. Ha sido bautizado como Protoceratops andrewsi, en su honor. Vuelva y encuentre más». Eso era exactamente lo que Roy Andrews intentaba hacer, precisamente, cuando la primavera volviera de nuevo a Mongolia. Andrews vio muchos cambios en la gran meseta cuando entró en Mongolia en abril de 1923. La vida esplendorosa había vuelto a lo largo de la ruta de la caravana de Kalgan a Urga. Las yurts se levantaban cerca de casi todos los manantiales, rebaños de cabras y ovejas pastaban en los pastizales del sur. Aquel año se unieron al tráfico algunos vehículos a motor, lo cual causaba una gran impresión en aquel paisaje desértico. Había transcurrido algo más de un año desde el cese de las hostilidades entre Mongolia y China y el país volvía a abrir sus puertas al comercio. La expedición de Andrews tomó la precaución de pasar la primera noche de camino en una posada china protegida por soldados y obtener una escolta militar fiable al cruzar parte de los territorios peligrosos. Pese a todas esas precauciones, algunos días más tarde, cuando Andrews con un pequeño grupo regresó a Kalgan para recoger parte del equipo que había llegado con retraso, tuvo un encuentro con tres bandidos a caballo. Escapó por los pelos lanzando su automóvil contra ellos, lo que hizo que sus caballos se espantaran y se encabritaran. La expedición puso rumbo a Iren Dabasu y volvieron a ocupar el mismo campamento en que estuvieron el año anterior, estableciendo una estación de recogida de fósiles. Por petición de Andrews, el Museo Americano les había asignado otros tres buscadores de fósiles que se incorporaron a la expedición del Asia Central. Se trataba de George Olsen y Peter C. Kaisen, que llevaban ya mucho tiempo trabajando en el museo, y Albert F. Johnson, que había trabajado con Bamum Brown en Alberta. Un joven chino que había formado parte del viaje de 1922, fue ascendido de criado y camarero a ayudante paleontólogo de Granger. Doce hombres podrían haber trabajado durante doce meses sin agotar los tesoros fósiles. Johnson explotó el más rico de los tajos, una fuente de huesos de dinosaurios carnívoros y herbívoros de distintas especies, muchos de los cuales eran dinosaurios de pico de pato. También encontró algunos fragmentos suavemente curvados, que recordaban las piezas de un rompecabezas perdido. Los Flaming Cliffs era el destino final de esta expedición. Siguiendo sus propias huellas de 1922, los hombres llegaron a Shabarakh Usu el 8 de julio y se quedaron allí cinco semanas. Los fósiles estaban por todas partes, pero el descubrimiento que daría fama mundial a la expedición ocurrió cinco días después, el 13 de julio.

Aquella tarde George Olsen regresó al campamento para informar de que había visto tres huevos fósiles en la piedra caliza. La piedra procedía del período cretáceo y, por lo tanto, probablemente era demasiado antigua como para contener huevos de grandes aves. Era posible que no fueran huevos sino cualquier tipo de formación geológica que había hecho que la roca adquiriera ese aspecto. Pero cuando Granger acompañó a Olsen al lugar del hallazgo e inspeccionó la cáscara estriada y marrón, llegó a la siguiente conclusión: «Tienen que ser huevos de dinosaurio. No pueden ser otra cosa». Los dinosaurios eran reptiles y éstos, con muy pocas excepciones, ponen huevos. El huevo de cáscara dura fue de hecho el invento evolutivo, conjuntamente con las escamas, gracias al cual los reptiles pudieron dejar el agua y pasar a tierra firme. Los primeros anfibios, como las ranas y las salamandras de nuestros días, tenían que llevar una doble vida. Podían salir del agua, pero tenían que volver con frecuencia al líquido elemento, no sólo para mantener húmeda su piel suave sino también para poner sus huevos desprovistos de cáscara dura. Los huevos normales de anfibio son partículas de embrión envueltas en gelatina, que se secarían muy rápidamente fuera del agua. Con el tiempo los reptiles descubrieron un modo de romper con los lazos que los ataban al agua. Fueron los pioneros en el desarrollo del huevo amniótico, una cápsula autocontenedora que podría ser puesta en cualquier parte. El embrión, que se desarrollaba dentro de la dura cáscara porosa, estaba rodeado por el líquido amniótico, una bolsa llena de albúmina como sustituto del agua y con la yema como reserva alimenticia. Este huevo, que los reptiles desarrollaron hace trescientos millones de años, en el período carbonífero, sirve como recordatorio de la ascendencia reptil de los mamíferos y las aves. Los humanos, como mamíferos, comparten este parentesco. Aunque nosotros no ponemos huevos, el óvulo humano, una vez fertilizado, queda envuelto en una bolsa de líquido amniótico muy parecido al del huevo de los reptiles. La envoltura del óvulo humano contiene también una pequeña bolsa de yema, pese a que el embrión es alimentado, a través de la placenta, por la sangre de la madre. Y el fluido en ese saco es salado. La evolución todavía tiene que erradicar este vestigio de nuestros orígenes en el mar, según el cual primero vino el pez que se arrastró fuera del agua a tierra firme, los anfibios que vivieron en ambos mundos, acuático y terrestre, y después los reptiles que con el advenimiento del huevo de cáscara dura estuvieron en condiciones de establecer su hogar permanente en tierra y dieron origen a los mamíferos. Por lo tanto no resultaba sorprendente descubrir que los dinosaurios habían sido animales ovíparos. La gran sorpresa hubiera sido descubrir que no ponían huevos. Pero hasta aquel momento nadie podía estar seguro de ese aspecto de la vida de los dinosaurios. Jamás se habían encontrado huevos fósiles intactos. Estaban aquellos trozos de cáscara encontrados en Flaming Cliffs en 1922, y en Iren Dabasu sólo unas pocas semanas antes, claves que nadie pudo entender hasta el hallazgo de Olsen.
En el campamento, los científicos y los recolectores estudiaron los tres huevos. Cada uno de ellos tenía unos veinte centímetros de longitud por diecisiete de circunferencia y eran algo más alargados y planos que los de los reptiles modernos. La cáscara tenía varios milímetros de grosor y su superficie externa era más bien arrugada y estriada mientras que la interior era lisa y suave. La expedición descubrió otros huevos, veinticinco en total, algunos agrupados en piña y otros en nidos. Granger encontró cinco juntos, formando un racimo. Johnson descubrió un grupo de nueve, dos de los cuales estaban rotos y exponían pequeños esqueletos de dinosaurios en estado de embrión. Una sección de roca que contenía varios huevos fue extraída y enviada en su totalidad al Museo Americano, donde se vio que contenía trece huevos en dos círculos. Sus puntos estaban dirigidos al centro tal y como debieron ser puestos en su nido arenoso decenas de millones de años antes para que fuesen incubados por el calor del sol. Pese a todo ese cuidado, por cualquier razón, aquellos huevos en particular nunca empollaron. Es posible que una ola repentina de frío impidiera la incubación o que una tormenta de arena los cubriera demasiado hasta el punto de impedir que llegara a ellos el necesario calor solar. En todo caso, los huevos se resquebrajaron, la arena fina entró en su interior y así comenzó el lento proceso de fosilización. Granger llegó a la conclusión de que los huevos encontrados en Flaming Cliffs eran del Protoceratops, el dinosaurio descubierto por Shakelford allí mismo en 1922. De los siete cráneos y catorce esqueletos recogidos por la expedición, allí, en 1923, muchos fueron identificados como restos de ese supuesto ascendente de los dinosaurios cornudos, oceratópsidos. Posiblemente Granger tenía razón. Pero también otros dinosaurios vivieron en Flaming Cliffs en aquellos tiempos. Debía de ser un lugar de puesta de huevos. Olsen encontró un pequeño esqueleto de dinosaurio que, cuando fue limpiado y puesto al descubierto, se vio que estaba agachado sobre un montón de huevos. Al parecer era un desdentado. Era posible que se alimentara sorbiendo los huevos del Protoceratops. Verdad o no, Osbom más tarde quedó impresionado por la prueba circunstancial al dar al animal en cuestión el nombre deOviraptor philoceratops. Oviraptor significa «capturador de huevos», y philoceratops significa «amante de los huevos de los ceratópsidos». Los nombres aplicados a los dinosaurios reflejan muchas veces más la imaginación que las pruebas reales. La posición del esqueleto sobre un nido de huevos, dijo Osbom, «hizo que de inmediato el animal resultara sospechoso de haber sido sorprendido por una tormenta de arena en pleno acto de robar un nido de huevos de dinosaurio».
A partir de otros huevos y cráneos los científicos identificaron a otro supuesto ladrón de huevos, el Velociraptor («ladrón rápido»), representado en distintas películas de la serie Jurasic Park; a un gran dinosaurio cubierto de escamas al que se le dio el nombre de Pinacosaurus, y al dinosaurio de aspecto de ave, el Sauromithoides. Este último tenía manos en forma de alas y un cráneo de constitución más bien ligera. Pero, como observó Andrews, era demasiado tarde en el tiempo geológico para ser un antepasado de las aves. Todos estos y muchos otros fósilesprotoceratopsianos eran nuevos para la paleontología. Al término de la expedición de 1923, Andrews regresó a Estados Unidos. Las noticias del hallazgo de los huevos le habían precedido y habían dado lugar a un torrente de publicidad en diarios y revistas. Un huevo es una cosa muy frágil y la idea de la existencia de huevos que fueron puestos por dinosaurios hacía millones de años y que habían sobrevivido en forma fósil, parecía algo increíble. Las fotografías de aquellos grandes objetos, tanto más que los huesos fósiles, documentaban la realidad de los dinosaurios como criaturas vivas que respiraron y se reprodujeron hacía muchos millones de años. Como truco publicitario se subastó uno de aquellos huevos que fue adquirido por Austin Colgate, tío de Bayard Colgate, que lo regaló a la Colgate University. Andrews lamentaría más tarde haber hecho aquella operación. Las autoridades chinas y mongoles al enterarse de la compra realizada por Colgate llegaron a la conclusión de que cada huevo de dinosaurio valía 5 000 dólares en los mercados mundiales y que si bien aquellos expedicionarios no habían ido a sus territorios en busca de petróleo o de oro sí buscaban algo igualmente valioso, aquellos huevos petrificados. Las siguientes expediciones fueron vigiladas con ojos cada vez más sospechosos. Andrews recibió la noticia, procedente de Nueva York, de que uno de los pequeños fósiles recogidos en Flaming Cliffs en 1923 era de un mamífero del período cretáceo, una rareza paleontológica de gran importancia. Hasta entonces sólo había sido descubierto un único cráneo de mamífero del mesozoico. Fue encontrado en África del Sur e identificado como perteneciente al grupo conocido como de los multituberculata, es decir, de unos mamíferos insectívoros de pequeño tamaño emparentados lejanamente con los modernos erizos y musarañas. Algunas de sus especies sobrevivieron a las extinciones que borraron a los dinosaurios de la faz de la tierra y siguieron existiendo durante algún tiempo más hasta que, finalmente, también llegó el final de su mundo. No dejaron descendientes directos, pero sus fósiles en las rocas arcillosas de Mongolia atestiguaban la existencia de abundantes especies de mamíferos entre los reptiles gigantes del mesozoico. En su carta dirigida a Andrews, Matthew escribió: «Haga todo lo que esté en sus manos para conseguir otros cráneos». Andrews dio instrucciones a Granger, que ya había regresado para excavar en Flaming Cliffs, y en una semana los hombres del equipo reunieron siete pequeños cráneos de mamíferos.
Los análisis subsiguientes demostraron que aquellos animales apenas si superaban el tamaño de una rata, tenían el cuerpo muy peludo y hocicos puntiagudos. No todos eran multituberculados, que formaban una rama colateral en la evolución de los mamíferos a partir de los reptiles. Los especímenes revelaron a William K. Gregory y a George Gaylord Simpson, paleontólogos del Museo Americano, que en la corriente principal del cretáceo los mamíferos, como clase zoológica, ya se habían separado de los marsupiales y los placentarios, esas especies en las cuales, después de dar a luz, la madre alimenta a sus fetos en la bolsa marsupial y aquellos otros que traían al mundo a sus hijos ya completamente formados. Esto confirmaba la anterior especulación de Huxley y Osbom de que los lejanos ascendientes de los mamíferos placentarios se podían situar en tiempos tan remotos como el período cretáceo, en forma de pequeños mamíferos, semejantes a los roedores, que se escabullían entre las patas de los poderosos dinosaurios. Andrews dirigió otras dos expediciones a Mongolia, en 1928 y 1930. Los científicos limitaron su campo de trabajo, en 1926, a la provincia de Yunnan. La guerra civil y la creciente xenofobia los mantuvo alejados de aquel escenario en 1927. Una de las expediciones de Andrews fue autorizada en 1928 y se descubrieron algunos otros restos de dinosaurios, así como algunos huevos, en Iren Dabasu, pero les fueron arrebatados por los soldados. Después de seis semanas de negociaciones, en los que intervino la embajada de los Estados Unidos que tuvo que recurrir a tan altas esferas como el ministro de Asuntos Exteriores de Nankín, se consiguió que los fósiles fueran devueltos a Andrews y a sus científicos. La última expedición, la quinta, se organizó en 1930, pero ésta no se adentró demasiado en Mongolia, pues para entonces las condiciones se habían hecho tan insoportables que Osbom dio instrucciones a Andrews para que cerrara el cuartel general de la expedición en Pekín y no hiciera ningún nuevo viaje a los hostiles lechos de fósiles de Mongolia. Los paleontólogos estaban empezando a revivir un pasado de interés más inmediato y directo para los seres humanos: nuestras propias raíces fósiles. En la década de 1850-1860 había sido descubierto el hombre de Neanderthal; el hombre de Java, el Homo erectus, en la de 1890-1900. Después, en 1924, Raymond Dart descubrió en África del Sur el llamado niño de Taung, el cráneo fósil del Australopithecus africanus. Ésta parecía ser la criatura que Darwin había predicho, a medio camino entre los antiguos simios y el hombre moderno. Andrews, actuando a espaldas de Osbom, había estado buscando en el lugar equivocado los orígenes del hombre. El descubrimiento de Taung puso en su lugar a la antropología y en las décadas siguientes el Australopithecus y el Ramapithecus se disputarían la atención del público con el Tyrannosaurus y el Brontosaurus. Lugares tales como el Gran Karoo, la garganta Olduvai y el valle Afar evocarían las historias del descubrimiento de los fósiles antaño asociados con Bernissart y Como Bluff, Hell Creek y Flaming Cliffs.
El modus operandi de la paleontología estaba cambiando. La mayor parte del mundo, con la excepción de las regiones polares, había sido recorrida y explorada ya por los cazadores en busca de fósiles vertebrados. En Europa como en Norteamérica, en África, América del Sur, en Australia y también en Asia, desde la India a través de China hasta regiones tan alejadas como Mongolia, habían sido hallados e identificados huesos de dinosaurio. Durante el período de los descubrimientos, la ciencia paleontológica inicia su investigación con la exploración a cielo abierto, a la que siguen el examen de los hallazgos, la acumulación de pruebas en un frente amplio, el seguimiento de las pistas que lleven a solucionar los puntos oscuros del problema y la elaboración de hipótesis bien fundamentadas. Los paleontólogos seguían saliendo al campo cada verano en busca de nuevos huesos. Aquellos nuevos exploradores del tiempo buscaron en los lechos jurásicos y cretáceos de América del Norte con un celo que no había vuelto a verse desde los días de Bamum Brown. Dinosaurio Jim Jensen, que había excavado los huesos del que bien podría haber sido el mayor de todos los dinosaurios, se convirtió en una celebridad menor para los niños. Los científicos encontraron un posible antepasado del Tyrannosaurus rexen el oeste de Texas. Se abrieron grandes yacimientos de dinosaurios fósiles en Alberta. Científicos polacos, rusos y mongoles volvieron a recorrer las zonas del Gobi antaño reconocidas por Andrews e hicieron descubrimientos espectaculares. La investigación se adentró más profundamente en el triásico. En 1984, un equipo dirigido por Robert Long, de la Universidad de California, descubrió en el Paitend Desert, Arizona, lo que se creyó podría ser el más antiguo de todos los dinosaurios conocidos hasta la fecha. Allí, en rocas que se calculó tenían doscientos veinticinco millones de años de edad, el equipo excavó el esqueleto de una criatura del tamaño de un avestruz pequeño. Algo más antigua que los Staurikosaurus descubiertos en América del Sur en la década de los sesenta, que anteriormente se habían considerado como los más antiguos de los dinosaurios, esa criatura podría muy bien representar una familia enteramente nueva de dinosaurios vegetarianos que podían estar emparentados con los antiguos Plateosaurus y que, tal vez, eran antepasados muy antiguos de los Brontosaurus. Pero el éxito de los cazadores de dinosaurios, en especial el de los profesionales, dejó de ser medido por el tonelaje de los huesos encontrados o por el número de nuevas especies que pudieron identificar. Los museos tenían ya tantos huesos que no sabían qué hacer con ellos, así que el interés principal se enfocó en la búsqueda de claves que descubrieran la anatomía, conducta y fisiología de los dinosaurios y dieran respuesta a los interrogantes. La nueva generación de paleontólogos empezó a preocuparse por estas respuestas y trató de situar el lugar de los dinosaurios a lo largo del tiempo.
Una figura en la transición de aquellos días fue Edwin H. Colbert. Comenzó a estudiar paleontología bajo la tutoría de Osbom y pasó a ser su ayudante investigador en 1930 y a ocupar el cargo de director de paleontología de los vertebrados en el Museo Americano. Escribió varios libros técnicos y de divulgación y, más recientemente, en 1983, uno tituladoDinosaurs: An Illustrated History. A él se le atribuye, con razón, el hallazgo de miles de fósiles, no siempre de dinosaurio, que incluyen más de cincuenta nuevas especies y diez nuevos géneros. En 1974 el Time llamó a Colbert «el Dick Tracy de la Era Mesozoica». Uno de sus descubrimientos más espectaculares lo hizo en el verano de 1947, en Ghost Ranch, cerca de Abiquiu, Nuevo México. George Whitaker, el ayudante de Colbert, dio con algunos huesos embutidos en las rocas de la pared de un barranco. Entre las primeras muestras recogidas resultó de especial importancia un pequeño trozo de una garra que Colbert identificó de inmediato como perteneciente a un Coelophysis. Entre los dinosaurios más antiguos de los conocidos hasta ahora, que vivieron hace más de 213 millones de años, el Coelophysis fue descrito por Cope en 1887, pero hasta ahora sus restos hallados han sido fragmentarios y escasos. Aquel verano Colbert recogió en Ghost Ranch los huesos de una docena de esqueletos completos. No se trataba de animales de gran tamaño, pues sólo medían un máximo de tres metros, y los huesos hablaban de la activa vida de carnívoros bípedos que debieron de vivir en los finales del triásico. Sus miembros posteriores eran alargados y recordaban a los de las aves, hechos para conseguir gran velocidad, y las manos tenían tres dedos terminados en garras dispuestos para dar caza a sus presas. Dentro de la cavidad corporal de al menos uno de los especímenes se encontraron huesos de un Coelophysis joven. Al principio Colbert mostró sorpresa por esto y tuvo sus dudas de que lo que estuviera viendo fuese un embrión de lo que hubiera sido la primera prueba de que algunos de los dinosaurios no eran ovíparos sino vivíparos, es decir, que traían vivos al mundo a sus hijos. La apertura de la pelvis, según observó, era demasiado pequeña y podía permitir el paso de huevos, pero no de animales vivos. Esto dejaba sólo una explicación alternativa. Colbert escribió: «La inevitable conclusión era que el Coelophysis fue un animal caníbal y en ocasiones se comía a sus propios hijos, exactamente como hacen algunos reptiles modernos. Una imagen no muy bella y agradable, pero sí realista». Puesto que resultaba imposible realizar experimentos fisiológicos con los dinosaurios, largo tiempo extinguidos, Colbert decidió que lo más útil que podía hacerse era investigar a los más parecidos entre sus parientes vivos, como los cocodrilos y caimanes. Con Raymond B. Cowles, un zoólogo de la Universidad de California, y Charles M. Bogert, también zoólogo, pero perteneciente al Museo Americano, Colbert capturó trece caimanes, algunos de los cuales llegaban a alcanzar los dos metros de largo, y los colocaron fuera del agua bajo el ardiente sol de Florida. Tomaron sus temperaturas a intervalos de diez minutos o menos. Cuando los caimanes alcanzaban temperaturas que probablemente hubieran causado su muerte, eran sacados del sol y conducidos a la sombra para que se enfriaran.

Los investigadores siguieron tomando sus temperaturas una y otra vez e, incluso, algunos de los animales fueron atados a marcos de madera para mantenerlos en posición erecta como la de los dinosaurios. Todo eso resultó excesivo para dos de estos animales, que murieron. El experimentó sirvió para que Colbert y sus colegas establecieran que el aumento y la disminución de la temperatura estaba en relación directa con la masa del animal. Los caimanes pequeños se calentaban más rápidamente al sol y se enfriaban más pronto a la sombra. Los mayores sufrían ascensos de la temperatura más lentos, e igualmente lo eran sus descensos. Colbert salió del experimento lleno de cardenales y pequeñas heridas por su lucha con los caimanes, pero con la impresión de que había encontrado una posible pista sobre el éxito de los dinosaurios. Aunque presumiblemente eran animales de sangre fría, como todos los demás reptiles, tenían la capacidad, gracias a sus grandes masas, de retener el calor corporal por períodos muy largos, lo que les daba la ventaja de la sangre caliente, es decir, de temperatura constante, que los mamíferos y otras criaturas de sangre caliente consiguen gracias a su elevado metabolismo. Así los dinosaurios podían ser activos como los mamíferos sin ser animales de sangre caliente. Los experimentos de Colbert con los caimanes fueron introducidos como una prueba de gran valor en los debates sobre si estos animales eran de sangre caliente a diferencia de los reptiles modernos. Colbert también participó en otro descubrimiento que aportaría la prueba decisiva en nombre de un nuevo concepto científico que no sólo explica la ubicuidad de los animales del mesozoico, como los dinosaurios, sino que ilumina la historia dinámica de toda la Tierra. No sería ésa la última vez que los intentos de resolver el enigma de los dinosaurios conducirían a sorprendentes teorías con respecto a la historia de la Tierra. En una expedición a la Antártida en 1969, Colbert, con Jim Jensen, encontró los huesos y el cráneo de un Lystrosaurus. Éste no era un dinosaurio sino una de las líneas más antiguas de reptiles que relevaron a los dinosaurios en la continua lucha por la supervivencia en la naturaleza. Era uno de los terápsidos, reptiles de aspecto de mamíferos que Robert Broom sacó al primer plano científico a finales del siglo pasado, en África. Colbert se había pasado algún tiempo en el Sur de África estudiando a losterápsidos, entre los que se incluía el Lystrosaurus, un animal robusto, de cuatro patas y del tamaño de un perro grande, así que ya sabía lo que buscaba en la Antártida. Si se encontraban en la Antártida los fósiles del triásico que ya habían sido encontrados en África esto podía considerarse como una prueba irrefutable de que Gondwana, el supercontinente del sur, había existido y, como respuesta a la presión de las placas tectónicas se había partido hacia finales del mesozoico.
Los científicos proclamaron el descubrimiento de Colbert-Jensen de un Lystrosaurus en otro continente como prueba irrefutable que señalaba la conexión de la Antártida con África, lo que daba mayor consistencia a la teoría de la deriva de los continentes. La interconexión de los continentes durante la mayor parte del mesozoico hacía más fácil entender por qué los dinosaurios, incluyendo muchas especies estrechamente emparentadas, existían en todo el mundo. Tan sólo en la Antártida no habían sido encontrados sus fósiles, hasta entonces, pero los científicos no se sentirían demasiado sorprendidos si un día, como ya ha ocurrido con elLytrosaurus, se encontraban allí fósiles de dinosaurios. La nueva generación de paleontólogos no sólo ampliaría y extendería los anteriores logros, como por ejemplo con el hallazgo de esqueletos completos de Coelophysis, o con la revisión del recientemente recapitadoBrontosaurus, sino que pasarían al campo más complejo y controvertido de la evolución del dinosaurio y de las razones y causas de su éxito y su fracaso. En la actualidad parece más importante que nunca, y también más posible, el entendimiento de los terápsidos. Estos animales dominaron la Tierra millones de años antes del auge de los dinosaurios. Tuvieron que ceder ante éstos y desaparecieron, pero no antes de que algunos de ellos sufrieran la trascendental transición de reptiles a los primeros mamíferos. Fueron los antepasados de todo gato o murciélago, ballena o primate, e incluso de todos los seres humanos. Menos espectaculares en apariencia que la mayoría de los dinosaurios, los terápsidos nunca desataron la imaginación humana del modo como lo hicieron los dinosaurios. En 1981 se celebró, por vez primera, una conferencia de importancia dedicada al estudio de su evolución. Pero el registro de fósiles de esos reptiles semejantes a los mamíferos y de sus antepasados es más completo que el de cualquier otro grupo de vertebrados terrestres, con la única excepción de los mamíferos del terciario. Fueron muy diversos y pasaron con éxito la evolución. Los antepasados de los terápsidos formaron el grupo de partida del género básico reptil hace ya más de trescientos millones de años, en el período carbonífero. Los reptiles acababan de desarrollarse de los anfibios. Una línea progresó en un sentido que podríamos llamar convencional, en dirección hacia los reptiles para acabar convirtiéndose en cocodrilos y dinosaurios, serpientes y lagartos. Otra línea, la subclase de los sinápsidos, de la clase de los reptiles, se convirtieron en reptiles con aspecto de mamíferos. Los primeros de ellos fueron los pelicosaurios, el más famoso de los cuales fue el Dimetrodon, dotado de una aleta dorsal como la del tiburón. Los terápsidosaparecieron en la Tierra hace unos 265 millones de años, al principio del pérmico, y hubieron de muchos tamaños y especies. Iban desde las dimensiones de una rata a las de un rinoceronte y, a veces, tenían formas ridículas.
John C. McLoughlin, en su libro Synapsida: A New Look into the Origen of Mammals, dijo: «Muchos de ellos nos hacen pensar que las fuerzas de la selección evolutiva hubieran sufrido un caso grave de delirium tremens durante el apogeo terápsido.» Algunos eran herbívoros, otros carnívoros y otros omnívoros. En época muy temprana comenzaron a mostrar signos de tendencias mamíferas. Pasaron de una postura horizontal semejante a la de los lagartos, a una forma de caminar algo más erecta, con las cuatro patas balanceándose bajo el cuerpo. El nombre de reptil, del latín reptilis, arrastrarse, empezó a convertirse en excesivamente restrictivo y pasó a ser, literalmente, como una descripción de los miembros extinguidos de esa amplia clase de animales. Aunque durante un siglo se supuso que los mamíferos descendían de los terápsidos, sólo hasta hace muy pocos años los científicos han tenido la certeza de esta vinculación. A partir de nuevos hallazgos de fósiles y de nuevas interpretaciones de anteriores descubrimientos, los científicos siguieron el rastro de los importantes pasos en la transición y determinaron que el momento decisivo debió de ocurrir a finales del triásico, hace unos 215 millones de años. Cráneos bien conservados de mamíferos de esa época, encontrados en el sur de China y en las reservas de los indios navajos en Arizona, Estados Unidos, resultaron pruebas muy importantes y reveladoras cuando fueron comparados con los cráneos de los terápsidos. En 1982, en su libro Mammallike Reptiles and the Origin of Mammals, Thomas S. Kemp, de la Universidad de Oxford, escribió: «Éste es un ejemplo conocido en el que la evolución de una clase de vertebrados procedente de otra clase está bien documentada por los fósiles». Fue en los cráneos, en particular en las mandíbulas, donde los científicos identificaron algunas de las más notables evidencias de que los terápsidos eran mamíferos en potencia, en tránsito evolutivo. Los fósiles terápsidos, encontrados en particular en África del Sur, en América del Sur y en Gran Bretaña, muestran el cambio que estaba teniendo lugar. Pero en los terápsidos no se encuentran algunas de las características que distinguen a los mamíferos de los reptiles. Tenían que desarrollar aún un cerebro mayor. De acuerdo con descubrimientos más recientes, elterápsido que parece más próximo a los antepasados directos de los mamíferos es un pequeño carnívoro conocido con el nombre de Probainognathus, cuyos fósiles se encontraron en los sedimentos del primitivo jurásico. En sus días de crepúsculo, sólo sobrevivían las más pequeñas especies de los terápsidos. Al parecer, los animales de mayor tamaño no sobrevivieron a la extinción masiva que se produjo a finales del período pérmico. Entre los más pequeños supervivientes emergió el Probainognathus y su descendiente o descendientes se desarrollaron hasta convertirse en, al menos, tres grupos de mamíferos primitivos: Morganucodon,Kuehneotherid y Amphilestid. El último, dice Crompton que parece «haber vivido felizmente durante un tiempo y desaparecer quién sabe dónde». Sin embargo, los descendientes delMorganucodon se desarrollaron en líneas que llevaron al Platypus y otros mamíferos actuales que son ovíparos. El Kuehneotherid se cree que es el ascendiente de casi todos los demás tipos de mamíferos.

En la época de la transición de los terápsidos a los mamíferos, a finales del triásico o comienzos del jurásico, los dinosaurios ya se habían autoestablecido como los herederos inmediatos de losterápsidos en el sentido de que en esos días eran los tetrápodos dominantes. Tetrápodo es el término científico que se emplea para los vertebrados terrestres y significa literalmente «cuadrúpedo». Los dinosaurios surgieron de otra rama importante de los reptiles. Aparentemente deben su auge, en parte, a la extinción masiva ocurrida en el pérmico y, también, parece como si la época de los dinosaurios estuviera encerrada entre los paréntesis de dos tragedias globales. Pronto, después de la extinción del pérmico, comenzaron a proliferar algunos reptiles que habitaban en los pantanos y fangales, los tecodontos. Tenían un aspecto parecido al de los modernos cocodrilos y un gran apetito por las restantes especies de losterápsidos. Con el tiempo los tecodontos pasaron y su existencia fue reemplazada, en el triásico, por los arcosaurios, los reptiles que reinarían durante el mesozoico. Ramas procedentes del tronco del tecodonto constituyeron cuatro grupos principales de arcosaurios,-los crocodílidos y los pterosaurios, así como dos líneas de dinosaurios, los saurisquianos y losomithisquianos. Entre los primeros omithisquianos se contaban los coelurosaurios, esos bípedos de cuerpo ligero y muy ágiles que representaba una notable diferencia en la norma de los reptiles. Uno de esos coelurosaurios eran el Coelophysis, cuyos huesos fueron encontrados de modo tan abundante por Edwin Colbert. Uno de los primeros saurisquianos, conocidos por elPlateosaurus, un animal de mucho mayor tamaño y un predecesor, en el triásico, delBrontosaurus y el Diplodocus, hallado, según se sabe, en las excavaciones realizadas en los años veinte por Friedrich von Huene, de la Universidad de Tubinga. Muchos de los terápsidosprobablemente no significaban un desafío para aquellos predadores ágiles e imponentes. Los que se alimentaban de día y dormían de noche eran especialmente vulnerables para los dinosaurios en auge. Los terápsidos que sobrevivieron durante el triásico, hasta que hicieron su transición a los mamíferos, debieron de haber encontrado algún nicho ecológico relativamente seguro. Por ejemplo, aquellos que evolucionaron mejoraron el sentido del oído y del olfato y podían, de algún modo, controlar la temperatura de sus cuerpos, rasgos peculiares de los mamíferos, y se convirtieron en criaturas nocturnas. Aquellos que no pudieron adaptarse a la vida nocturna dejaron de existir. Por esa razón sobrevivieron aquellos terápsidos que tenían más características propias de los mamíferos y transmitieron sus genes a sus descendientes que se fueron haciendo cada vez más mamíferos hasta terminar siéndolo. El legado de su estrategia de supervivencia se prolongó durante una gran parte del mesozoico. Los pequeños mamíferos que vivían a la sombra de los poderosos dinosaurios eran, generalmente, criaturas nocturnas.
Al comparar los terápsidos y los primitivos mamíferos con los dinosaurios, Robert T. Bakker define como divinas las razones de la «superioridad» de los dinosaurios en el mesozoico. Por lo corriente sólo se han conservado en estado fósil las partes duras (huesos y dientes) de los animales vertebrados. Sin embargo, músculos, tendones, nervios y vasos sanguíneos, y en ocasiones algunos otros tejidos y órganos blandos dejaron su marca en la superficie de los huesos. Detallados análisis de la anatomía de diversos vertebrados vivos revelan cómo esos órganos blandos afectan las superficies de los huesos. A partir de esas interpretaciones se hace posible deducir algunas cosas de la actividad, fisiología e, incluso, de la conducta de animales que ahora nos son conocidos sólo por sus huesos fosilizados. Ese nuevo método de estudiar los vertebrados fósiles está aportando algunas pruebas que desafían muchas de las teorías sobre los hábitos y la ecología de uno de los grupos más populares de animales extinguidos, los dinosaurios. Esas grandes bestias eran reptiles cuyos parientes más cercanos aún vivos son los modernos cocodrilos. Generalmente, los paleontólogos aceptaron que en todos los detalles de su vida, los dinosaurios eran simplemente cocodrilos, lagartos o caimanes que habían crecido más de lo normal. Los crocodílidos y los lagártidos pasan gran parte de su tiempo inactivos, tomando el sol echados en la roca o el tronco más conveniente, y comparados con otros animales modernos, la mayor parte de los reptiles modernos son lentos y perezosos. De aquí viene que la reconstrucción de un dinosaurio como el Brontosaurus nos lo presente como una montaña de carne fláccida que se mueve a su alrededor sólo de forma lenta y poco frecuente. Esta visión del dinosaurio, señalaba Bakker, presentaba un «problema que sumía en la perplejidad». ¿Cómo fue posible que estos lentos y perezosos dinosaurios dominaran a losterápsidos, que se estaban desarrollando activamente en la línea hacia los mamíferos y pudieron mantener sometidos a los mamíferos a lo largo del mesozoico? Bakker continuó mostrando evidencias basadas en las posturas y en los sistemas cardiovasculares de los dinosaurios que demostraban que aquellos animales fueron «criaturas ágiles, rápidas y llenas de energía y que vivían a un nivel fisiológico alto, sólo conseguido en todas partes entre los vertebrados terrestres por los mamíferos últimos y más avanzados». Para ilustrar esta nueva visión del dinosaurio, Bakker dibujó para la primera página de la revista un boceto de una pareja de dinosaurios cornudos al galope, y dijo que eran capaces de alcanzar una velocidad de hasta cincuenta kilómetros por hora. Al parecer había más vivacidad en los dinosaurios de la que los científicos les habían atribuido o imaginado corrientemente. En la conclusión Bakker escribió: “Ahora podemos comenzar a responder a la cuestión planteada al principio de este artículo: ¿por qué los reptiles con rasgos mamíferos perdieron en su competencia con los dinosaurios?”
Al parecer los mamíferos no adquirieron la forma más eficaz de locomoción erecta hasta bien entrado el período cretáceo, a finales de éste, es decir, cien millones de años después de que la lograran el evolucionado tecodonto y los primeros entre los dinosaurios. En la actualidad, al pensar en los mamíferos nos los presentamos como criaturas ágiles, activas y llenas de energía mientras que a los reptiles los vemos como animales que, de acuerdo con su nombre, se arrastran, reptan, lentamente por el suelo. Sin embargo, lo cierto es que los dinosaurios y sus parientes consiguieron notables avances en su locomoción mucho antes de que los lograran los mamíferos, y la superioridad de sus extremidades fue sin duda el factor más importante en el éxito del arcosaurio y la extinción de los reptiles con características de mamífero. Ya bien entrada la noche, en un día de agosto de 1964, John Ostrom y su ayudante Grant E. Meyer recorrían una ladera en el sur de la Montana central, cerca de la ciudad de Bridger. Se trataba de una pradera, campos de yerba interrumpidos por lugares erosionados y situados entre colinas de pinos y enebros. Ostrom y Meyer acababan de examinar un lugar que pensaban convertir en sede de excavaciones al año siguiente e iban a lo largo de la ladera, de un terraplén erosionado a otro; se habían alejado ya unos setecientos metros del lugar elegido cuando vieron unas garras que sobresalían de la tierra. Aquellos huesos habían estado allí años y años sin ser reconocidos. En cuestión de minutos pusieron al descubierto otras partes de aquella mano. Los diversos huesos de los dedos tenían la longitud aproximada de los dedos de un ser humano adulto. Las garras eran largas y afiladas. Ostrom y Meyer habían encontrado una poderosa mano con tres dedos prensiles. También hallaron algunos dientes, los dientes afilados, de sierra, de lo que fuera un carnívoro. Al día siguiente regresaron provistos de otras herramientas más útiles que sus navajas, y los dos paleontólogos continuaron excavando hasta hacer un descubrimiento aún más excitante: los huesos perfectamente conservados de un pie. Se lo quedaron mirando llenos de sorpresa. En todos los demás dinosaurios carnívoros los pies traseros son, por lo general, de forma parecida a los de las aves, con tres dedos principales y uno más pequeño en la parte interior o en la posterior del pie. Por lo general, esos tres dedos principales son iguales de forma, con el central algo más largo y los otros dos iguales entre sí en longitud y divergentes del central. Es decir, en conjuntos semejantes a los de las aves. Pero el pie de la criatura que ellos habían encontrado difería del plan básico general. El dedo externo y el medio tenían la misma longitud y el más interno de los dedos principales salía hacia fuera «como un pulgar hinchado», dijo Ostrom. Este dedo interior era más largo que los demás y en lugar de tener una corta garra puntiaguda y triangular como los otros, estaba dotado de una garra larga y delgada con la forma curvada de una hoz.

Ostrom jamás había visto antes nada comparable a aquella terrible garra, pero podía imaginarse bien el uso que el animal debió de hacer de aquel instrumento. Para aquella criatura que vivió hace más de ciento veinticinco millones de años, Ostrom pensó el nombre de Deinonychus, que significa «garra terrible». El Deinonychus es el más notable de los dinosaurios hasta ahora descubiertos. Casi todo lo relacionado con ese animal, sus brazos y piernas, sus terribles garras y su cola rígida, fue pronto presentado como prueba de la existencia de los dinosaurios rápidos, ágiles y dinámicos de Bakker. La evidencia, según la vieron muchos paleontólogos, parecía abrumadora, pero el debate subsiguiente polarizaría la paleontología de los dinosaurios. ¿Formaban éstos un grupo de animales de sangre caliente como las aves y los mamíferos? ¿Era su metabolismo más parecido al de los mamíferos que al de los reptiles, lo que les daba capacidad para vivir una existencia más activa? Ostrom, en su examen inicial del Deinonychus, comenzó a sospechar que ése podía ser el caso. Pero la idea, en sí, contradecía el punto de vista tradicional que presenta a los dinosaurios como criaturas tan perezosas y lentas como los reptiles, basándose en su anatomía ósea, que parece indicar que debieron de ser animales de sangre fría. Esta deducción lógica no fue discutida seriamente hasta que una nueva generación de paleontólogos empezó a contemplar a los dinosaurios, en especial al Deinonychus, bajo un nuevo ángulo. Durante un período de tiempo demasiado largo los paleontólogos parecieron ciegos con respecto a los atributos no reptiles de los dinosaurios. Sólo porque un determinado animal fósil sea integrado en una clase particular no debe deducirse que posee todas las características y atributos de los miembros modernos de esa clase. Al examinar los esqueletos, Ostrom llegó a la conclusión de que el Deinonychus era relativamente pequeño, en relación con otros dinosaurios y, además, también más ligero. Tenía una altura de un metro y medio aproximadamente y desde el hocico hasta la punta de la cola debía de medir dos metros y medio. A juzgar por los huesos de sus extremidades y por sus vértebras, el animal en plenitud de su crecimiento no debía de pesar más de ochenta kilos. A juzgar por sus dientes era un carnívoro, y así fue clasificado con los dinosaurios saurisquianos, el mismo suborden que losTyrannosaurus. La estructura de las extremidades delanteras y de las manos mostraba claramente que el animal era forzosamente un bípedo y que no podía, en ningún caso, marchar a cuatro patas aunque hubiese querido hacerlo. Al igual que muchos otros reptiles, elDeinonychus tenía una cola larga en relación con su longitud total, aproximadamente la mitad de ésta, pero, además, la cola terna una característica totalmente distinta a todo lo que Ostrom había visto hasta entonces. En su entera longitud la cola estaba como encajada en unos canutos óseos y paralelos, que no eran más que tendones osificados, como los que Dollo había hallado en los Iguanodontes belgas. Al principio, esto extrañó y llevó a error a Ostrom, hasta que se dio cuenta de que una cola rígida, que presumiblemente resultaba posible debido a las «varillas» osificadas, debía de haber sido muy importante para el Deinonychus.
Tal y como Ostrom se la representaba, aquella cola rígida podía ser movida hacia arriba y hacia abajo, hacia los lados y girando como las manecillas de un reloj. La cola, concluyó, era utilizada como un estabilizador dinámico, una especie de contrabalance o de equilibrador necesario para un animal bípedo muy activo y móvil. Eso, y otras evidencias halladas en el esqueleto, le indicaban a Ostrom que la postura de aquel bípedo era mucho más parecida a la de un avestruz, con el tronco mantenido casi en posición horizontal, el cuello curvado hacia arriba y la cola surgiendo recta por detrás. «En mi opinión -escribió Ostrom en 1969-, ésta resultaba una postura de aspecto mucho más natural que la postura del canguro, con la que se suele ilustrar generalmente a otros dinosaurios carnívoros, como el Allosaurus o el Tyrannosaurus». Añadió que si el esqueleto de esos gigantes hubiese estado tan bien preservado como los delDeinonychus, los científicos probablemente hubiesen podido observar que también ellos tenían con frecuencia sus colas levantadas y las utilizaban como contraequilibrio rígido, más que dejarlas arrastrar por el barro. Si el Deinonychus andaba y corría sobre sus dos extremidades traseras esto hacía más notable aún la presencia de la terrible garra, una especie de espolón curvo, en cada pie. Podía esperarse esa especie de garra en una mano prensil, razonó Ostrom, pero no en el dedo de un pie que está en contacto con el suelo, lo cual hacía muy posible que se rompiera o despuntara, incluso que hiriera a su propio dueño y lo dejara indefenso. Cada una de esas garras sobresalía unos diez o doce centímetros, ciertamente un arma defensiva de gran eficacia que, posiblemente, también podía ser utilizada para dar muerte y descuartizar a sus presas. «El sentido común nos dice que una hoja curvada como una hoz, afilada y fina sirve para cortar o desgarrar y no para excavar, subirse a los árboles o facilitar el desplazamiento en tierra», dijo Ostrom. Pero ¿cómo manejaba el animal esa arma tan letal? ¿Cómo la protegía cuando corría? A esta última cuestión los fósiles parecían ofrecer una respuesta directa. Las dos principales articulaciones del dedo interno, el que poseía la garra en cuestión, resultaban excepcionales en el sentido de que permitían que la garra pudiera ser alzada y desplazada hacia arriba y hacia atrás, con lo que se lograba separarla del suelo. Las articulaciones de los otros dedos de las patas traseras no eran retráctiles. La respuesta a la otra pregunta, la de cómo utilizaba el animal la garra, llevó a Ostrom a interpretaciones que señalan la importancia delDeinonychus para los paleontólogos. «A nadie nos sorprende ver cómo un águila o un halcón golpean con sus espolones al saltar o se mantienen de pie sobre una pata y golpean con el espolón de la otra -escribió Ostrom -, pero resulta ridículo imaginarse a un lagarto o a un cocodrilo (o cualquier otro de los modernos reptiles) irguiéndose sobre sus patas traseras para lanzarse al ataque, simplemente porque los reptiles son incapaces de tan complicadas maniobras, de esos juegos de equilibrio tan delicados, por falta de agilidad, como también son incapaces de una actividad tan exigente metabólicamente. Como sabemos, los reptiles son animales lentos y perezosos que se arrastran o permanecen inactivos la mayor parte del tiempo».
La garra en forma de hoja de hoz requería del Deinonychus que hiciera precisamente lo mismo que el águila oel halcón, es decir, que atacara con uno o con ambos pies a la vez. Eso no tiene nada que ver con el andar o el correr o el permanecer de pie, pero podía explicar la utilidad de la cola como elemento equilibrador. Sugiere, además, la existencia de una bioenergía impropia de un reptil que resultaba necesaria para poder mantener una lucha de ese tipo. Ostrom llegó a la siguiente conclusión: «Esa criatura debía de estar de pie o saltar sobre un pie u otro mientras golpeaba con el opuesto. Tal equilibrio y agilidad son desconocidos en cualquiera de los actuales reptiles vivos». El examen de los brazos y de sus largas manos prensiles parecen haber reforzado la opinión de Ostrom de que el Deinonychus era un depredador inteligente. La articulación de la muñeca giraba para permitir que las manos se volvieran una hacia la otra. De ese modo el animal podía sujetar la presa con las manos y trabajar con ambas en conjunción. Eso es algo que sólo pueden hacer los seres humanos y algunos otros mamíferos. ElDeinonychus y algunas especies emparentadas con él eran los únicos dinosaurios que, por lo que se sabe, tuvieron esa especial movilidad de las muñecas. La resonancia de los informes de Ostrom de 1969 sobre el Deinonychus y el metabolismo de los dinosaurios aún continúa teniendo eco en los estudios sobre el dinosaurio. En un informe en el Bulletin del Museo Peabody, Ostrom resume las consecuencias que pudo deducir de los esqueletos que había descubierto en Montana. Escribió: «El pie del Deinonychus es, posiblemente, la prueba más reveladora, desde el punto de vista anatómico, de los hábitos de los dinosaurios en el sentido de que éstos debieron de ser cualquier cosa, pero no “reptiles” en su conducta, en sus reacciones y en su forma de vida. Este dinosaurio tuvo que ser un animal de carrera rápida, altamente predador, extremadamente ágil y muy activo, sensible a muchos estímulos y rápido en sus reacciones de respuesta. Esto, a su vez, indica un nivel de actividad poco corriente para un reptil y sugiere la existencia de un ritmo metabólico alto. Las pruebas en favor de esa teoría radican principalmente, aunque no de modo exclusivo, en el pie». Es bien sabido, observó Ostrom, que los peces, los anfibios y los reptiles modernos son lo que se ha dado en llamar animales de sangre fría. El término «sangre fría» significa que la temperatura del animal fluctúa de acuerdo con la del ambiente que lo rodea. El animal de sangre fría carece de un mecanismo interno que le permita elevar o descender su temperatura muy por encima o por debajo de la ambiental. La mayor parte de los reptiles están inactivos en el frío de la noche y tienen que esperar a que salga el sol para calentarse y conseguir energías para la caza cotidiana. Puesto que su principal fuente de calor es externa, los reptiles y otros de los llamados animales de sangre fría son ectotermos. Cuando los ectotermos comienzan a estar demasiado calientes, tienen que buscar la sombra. Siempre dependen de la temperatura y deben actuar de acuerdo a sus fluctuaciones. En contraste, los animales de sangre caliente, mamíferos y aves, tienen mecanismos internos de regulación de la temperatura corporal, que mantienen aproximadamente al mismo nivel, con independencia de las condiciones externas. Estos animales son conocidos como endotermos.

Los seres humanos, salvo en casos de enfermedad, mantienen una temperatura de 37,1 grados centígrados. La mayor parte de los otros mamíferos operan a temperaturas comparables, mientras que las aves tienen, por lo general, temperaturas algo más elevadas, en torno a los 40 grados centígrados. Muchos endotermos sudan y jadean para ayudarse a rebajar la temperatura y mantenerse frescos o tienen capas de pelo o de plumas para evitar la pérdida del calor corporal. Pero el mecanismo principal de la endotermia es un metabolismo básico elevado. Todas las células vivas generan una pequeña cantidad de calor como producto secundario de los procesos químicos que se dan en su interior, y en los mamíferos y las aves ese proceso, conocido como metabolismo, es al menos cuatro veces superior en actividad a los ectotermos de cuerpo y temperaturas comparables. Como consecuencia los endotermos pueden ponerse en acción en casi cualquier condición climatológica, mientras que los ectotermos están gravemente influenciados y limitados por las condiciones ambientales que los rodean. Es esta dependencia climática de los ectotermos, dijo Ostrom, la que hace a estos animales muy útiles para la interpretación paleoclimatológica. Si los dinosaurios eran ectotermos, de acuerdo con las anteriores presunciones, entonces los millones de años durante los cuales vivieron debieron de ser tiempos de un clima suave a extensión mundial. Sólo en un medio ambiente suave y equilibrado, sin grandes altibajos, esos reptiles pudieron haber prosperado en tal número y en lugares que al parecer se extendían por toda la Tierra, hasta el límite de las regiones polares. Se puede presumir que no hubieran soportado cambios estacionales extremos. Sin embargo, aunque el hallazgo de polen y de fósiles invertebrados tiende a apoyar la idea de que el mesozoico se caracterizó por una climatología suave, Ostrom arguye que los paleontólogos no deben recurrir a los dinosaurios para corroborar esa evidencia. Afirmó que los dinosaurios son inútiles como indicador termal, porque hay motivos para cuestionar la presunción de que su metabolismo era reptiliano, es decir, su ectotermia. Ostrom declaró: «Hay pruebas considerables e impresionantes, si no decisivas, de que muchos tipos de los antiguos reptiles se caracterizaban por poseer un nivel de metabolismo comparable al de los mamíferos o las aves». El Deinonychus y, aparentemente, muchos otros dinosaurios, al igual que hacen los bípedos o los cuadrúpedos, se mantenían erectos con los pies directamente bajo el cuerpo y no extendidos y despatarrados por fuera, como es característico en los reptiles actuales. Ningún ectotermo vivo tiene una postura semejante. Si fuera así estarían en condiciones de correr con mayor rapidez y recorrer mayores distancias de lo que lo hacen. Al menos algunos dinosaurios eran ligeros de pies. Las largas extremidades del Deinonychus, y de algunos otros dinosaurios, a deducir de lo que indican sus fósiles, fueron rápidos corredores. «La evidencia parece indicar -dijo Ostrom- que la postura y la locomoción erectas probablemente no son posibles sin un metabolismo elevado y una alta y uniforme temperatura corporal».
Con ello no afirma claramente que los dinosaurios fueran animales endotermos. Podían haber sido, también, homeotermos, es decir, capaces de mantener una temperatura corporal constante por cualquier otro método, externo o interno. Tanto si eran homeotermos como, posiblemente, endotermos, afirma Ostrom, lo cierto es que los dinosaurios fueron unos animales extremadamente activos, cuyos mecanismos bioenergéticos los apartaban, al parecer, del mundo de los reptiles corrientes. Muchos paleontólogos encontraron inaceptable la idea de que los dinosaurios fuesen animales de sangre caliente. Otros, por el contrario, la encontraron liberadora, puesto que ofrecía un modelo de fisiología del dinosaurio que podía explicar su prolongado éxito y, posiblemente, incluso su extinción. La reconstrucción de la fisiología térmica de los animales extinguidos no es asunto fácil. En los fósiles no hay nada que aporte información sobre cómo regulaban los dinosaurios la temperatura de sus cuerpos. Todas las pruebas aportadas en favor o en contra de la hipótesis endotérmica están derivadas de las deducciones científicas extraídas de los huesos de los dinosaurios. Los resultados han sido sorprendentes. Distintas interpretaciones de los mismos huesos producen hipótesis y suposiciones en conflicto. La observación microscópica de los huesos del dinosaurio aporta pruebas que parecen apoyar las ideas de Ostrom. Ya en 1957 los paleontólogos se mostraron extrañados cuando encontraron en los huesos de los dinosaurios una red de delgados vasos sanguíneos (conductos de Havers) penetrantes destinados a aportar al tejido óseo cantidad abundante de sangre rica en nutrientes. Estos huesos con una red de delgados vasos sanguíneos son capaces de un crecimiento rápido y son indicadores, además, de la existencia de un nivel de metabolismo elevado. En 1968, Armand de Ricqles, un anatomista y paleontólogo de la Universidad de París, comenzó a informar de los resultados de estudios más detallados de los tejidos óseos y llegó a la conclusión de que aquel denso tejido de conductos de Havers indicaba «niveles de intercambio de fluidos hueso-cuerpo que, al menos, se aproximaban al de los grandes mamíferos vivos en la actualidad». Esto, en principio, parece ser la prueba más directa que sugiere, como dice Ricqles, «altos niveles de metabolismo y, en consecuencia, la endotermia entre los dinosaurios».
Ostrom continuó basando sus hipótesis en el hecho de que la postura erecta y el modo de andar que de ella se deriva sólo se da en animales endotérmicos, mamíferos y aves. Pero reconoce que las críticas que se le hacen tienen un punto de razón cuando arguyen que no existe una relación de causa-efecto entre la postura y la fisiología, ni nunca fue establecida. Sin embargo, insiste, «la correlación entre postura y endotermia o ectotermia es, virtualmente, absoluta y seguramente no se trata de una mera coincidencia». Adelanta otra línea de evidencia indirecta relacionada con la postura erguida. Citando las investigaciones de Roger S. Seymour, un zoólogo de la Universidad de Adelaida, en Australia, Ostrom observa que la mayor distancia vertical entre el corazón y el cerebro de un animal requiere una presión sanguínea mayor. La presión sanguínea de la jirafa es doble de la del ser humano. En el caso delBrachiosaurus, para tomar un ejemplo extremo entre los dinosaurios, la distancia entre el corazón y el cerebro era de aproximadamente seis metros. Para bombear sangre a esa distancia, e incluso a otras notablemente menores, en un animal con postura erguida, dice Ostrom, se requeriría un corazón de cuatro ventrículos, muy avanzado, lo que constituye una característica de los animales endotérmicos. Owen, en 1841, se preguntó si no era posible que los dinosaurios hubieran tenido un corazón de cuatro ventrículos, pero no siguió investigando en este sentido. Los cocodrilos tienen una versión imperfecta de este corazón de cuatro ventrículos, lo que parece indicar que no puede desecharse esa versión en los saurios primitivos. La distancia cerebro-corazón no prueba que los dinosaurios fueran endotermos, como observa Seymour, pero ayuda a pensarlo así el hecho de que muchos de ellos tenían necesidad de un corazón y un sistema circulatorio capaz de mantener una fisiología endotérmica. En contraste con las precavidas deducciones de Ostrom y sus concesiones a la crítica, un antiguo discípula, Robert Bakker, afirmó que la hipótesis en favor de la endotermia de los dinosaurios era decisivo. Los animales de sangre caliente pagan un alto precio por el metabolismo que los mantiene en un estado constante de disposición a la acción. Tienen que comer más y eso significa tener que pasarse más tiempo pastando o cazando. Un león consume su peso en alimentos cada siete o diez días, mientras que el dragón de Komodo, un lagarto carnívoro, come su peso en alimentos sólo cada sesenta días. Esto significa que una determinada cantidad de carne podrá alimentar a un mayor número de carnívoros ectotérmicos que endotérmicos. Consecuentemente Bakker estableció la relación entre las poblaciones de presas y de carnívoros depredadores, tal y como se mostraba en los fósiles de finales del cretáceo. Examinando varias colecciones de fósiles, separando los predadores de sus presas por los dientes y maxilares y calculando el probable peso del cuerpo, Bakker intentó determinar el porcentaje de predadores en la totalidad de población fósil.

Un pequeño porcentaje de predadores significaría que habían tenido un buen apetito endotérmico. En la actualidad, en algunos hábitats africanos, los predadores constituyen tan sólo entre el uno y el seis por ciento de la población animal en su totalidad. Para los dinosaurios, a deducir de las muestras fósiles, Bakker encontró que los predadores representaban entre el uno y tres por ciento del total. La relación presa-predador de los reptiles premesozoicos, que eran indiscutiblemente ectotérmicos, alcanzaba entre el treinta y cinco y el sesenta por ciento. Esos datos se convirtieron en la base fundamental de la campaña, cada vez más agresiva, de Bakker, en favor de la endotermia de los dinosaurios. Presentó su defensa de la endotermia de los dinosaurios en un artículo publicado en Scientific American que tituló «Dinosaur Renaissance» («El renacimiento del dinosaurio»), en el cual afirmó: «Las relaciones predador-presa son instrumentos muy poderosos y útiles para la paleofisiología porque son los resultados directos del metabolismo de los depredadores». Mientras tanto, Ostrom había modificado su postura en relación con la endotermia de los dinosaurios. Bakker necesitaba establecer la endotermia de los dinosaurios para reforzar su teoría que explicaba la ascendencia de los dinosaurios sobre los mamíferos en el mesozoico, suspendiendo su evolución en tanto que estos últimos tuvieron que competir con los dinosaurios. En la mente de Bakker sólo animales endotérmicos muy activos podían mantener la «superioridad» de los dinosaurios en el mesozoico. Ostrom no había cambiado su opinión sobre el ágil Deinonychus o sobre la vitalidad de muchos otros dinosaurios; la garra, la cola y su locomoción erecta y bípeda parecían seguir siendo una prueba evidente e indiscutible en favor de su gran nivel de actividad. Pero Ostrom no podía ir tan lejos, basándose sólo en esas pruebas, como para unirse a Bakker en sus arrasadoras generalizaciones sobre la biogenética de los dinosaurios y su superioridad. «Si Bakker se hubiese movido con mayor cautela, hubiéramos encontrado menos ardor y una mejor acogida -diría Ostrom en 1983-. Nunca me fue posible convencerle de que la exposición mesurada siempre es superior a la exposición exagerada». Los argumentos en favor y en contra de la endotermia se airearon en la reunión anual de la Asociación Americana para el Avance de la Ciencia, en Washington. Los partidarios de la endotermia estaban encabezados por Bakker y, en menor grado, por Ostrom y Ricqles. Nicholas Hotton III, de la Institución Smithsoniana, surgió como líder de la oposición, aunque tenía importantes aliados en Dale Russell y Philip Regal de la Universidad de Minnesota y James Spotila de la Universidad del Estado de Nueva York, en Buffalo. Los resultados del simposio, publicados en 1980 como libro bajo el título A Coid Look at the Warm-Blooded Dinosaurs, aparecieron como un documento definitivo en la controversia.
Uno de los argumentos contra Bakker se referían al gran tamaño de los dinosaurios. Éstos se comportaban posiblemente como animales de sangre caliente no porque fuesen endotérmicos sino porque eran muy grandes. Es posible que debieran su éxito a su extraordinario tamaño, a su enorme masa. Si hay algo en relación con los dinosaurios que está por encima de toda disputa es su tamaño. Hotton determinó que el ochenta por ciento de los mamíferos vivos son más pequeños que el más diminuto de los dinosaurios, que pesaban unos diez kilogramos, y que más de la mitad de los dinosaurios pesaban más de dos toneladas, un peso alcanzado tan sólo por el dos por ciento de los actuales mamíferos. Parece ser, pues, que el gran tamaño fue, en cierto modo, un factor crítico para la supervivencia del dinosaurio, y los individuos mayores de una especie son los más aptos para reproducirse conduciendo a la evolución de especies de animales aún mayores, al menos hasta alcanzar cierto punto en el que las ventajas disminuyen. Algunos científicos presentaron datos sugiriendo que los ectotermos y los endotermos se hacen más parecidos entre sí a medida que aumenta su tamaño. Los requerimientos metabólicos y consecuentemente la relación depredador-presa podrían ser muy semejantes si ambos son grandes. Según defiende Spotila, un cuerpo con una gran masa produce una temperatura corporal bastante constante. Los dinosaurios podrían haber sido animales inactivos homeotérmicos capaces de mantener una temperatura corporal constante por cualquier medio, incluyendo aquellos dependientes del medio ambiente. Colbert y sus colegas en sus primeros experimentos con los caimanes de Florida, habían demostrado que esos animales, descendientes de los arcosaurios, se calientan más lentamente cuando son expuestos a la luz solar y también se enfrían más lentamente que otros animales más pequeños. Estos caimanes pueden mantener las temperaturas de sus cuerpos relativamente constantes sin necesidad de un mecanismo endotérmico o sin un aislamiento exterior, como las pieles peludas o las plumas. Cuanto mayores son los caimanes, descubrió Colbert, más lento es el ritmo de absorción y de pérdida de calor. En el clima cálido del mesozoico las variaciones de la temperatura corporal de los animales podrían ser presumiblemente mínimas. Por consecuencia, los dinosaurios podrían tener los atributos de los animales de sangre caliente sin necesidad de ser endotérmicos.
Hotton cree que fue la homeotermia del dinosaurio la clave: «Los dinosaurios, al igual que los mamíferos, aumentan su capacidad de actividad continuada reduciendo su dependencia del medio ambiente físico como fuente de calor corporal. Ello lo hicieron así a bajo coste, mediante un sistema de conservación de calor que configura un estilo de vida que fue muy diferente de los estilos de vida de otros animales, condicionados por el sistema de generación de calor de los mamíferos que requiere un alto coste. Los mecanismos locomotores y el tamaño ilustran una diferencia fundamental: la actividad de los dinosaurios era más tranquila que la de los mamíferos. En general la estrategia básica de los dinosaurios era “lenta y utilitaria” y lo que perdía en impulso en relación con los mamíferos lo ganaba en economía». Una parte de la estrategia, sugirió Hotton, involucra probablemente migraciones estacionales sobre distancias que sobrepasaban los 3.200 kilómetros. En Norteamérica se han encontrado fósiles de dinosaurios en latitudes tan nórdicas como el Territorio del Yukón, a sesenta grados de latitud norte. Incluso en el suave clima del mesozoico, el Ártico no era probablemente el lugar más adecuado para los dinosaurios, sobre todo en invierno, debido al frío y a la oscuridad que interrumpían el crecimiento de las plantas. Los grandes dinosaurios, tanto herbívoros como carnívoros, tuvieron que emigrar para poder conseguir suficiente alimento para mantenerse en acción, incluso viviendo al modesto ritmo del metabolismo ectotérmico. Hotton ofreció otras dos razones más para explicar esas migraciones. La primera era que la propia actividad migratoria podría facilitar una fuente de calor interno en la que se podía confiar; la segunda, que los viajes los habrían mantenido expuestos aproximadamente a la misma temperatura durante todo el año, lo que significaba una consideración vital si, en su calidad de animales ectotérmicos, la tolerancia de los dinosaurios a las fluctuaciones térmicas estaba limitada dentro de unos márgenes relativamente pequeños de emisión de calor de un metabolismo en descanso. El concepto de que algunos dinosaurios, como muchas aves que aún viven en la actualidad, emigraran al norte o al sur, estación tras estación, es algo sorprendente pero no imposible de creer. Hotton también participaba de la opinión de Bakker de que la endotermia podía haber sido un factor que, conjuntamente con el gran tamaño y la piel desnuda de los dinosaurios, participara en las causas de su extinción. Bakker había afirmado: «Frente a un estrés causado por un frío repentino y prolongado, los dinosaurios eran demasiado voluminosos para escapar hibernando en madrigueras o en otros microhabitáculos que estaban a disposición de los demás animales endotérmicos, y, por otra parte, eran incapaces de sobrevivir a prolongados descensos de su temperatura corporal, como ocurre con algunos tipos de tortugas, lagártidos y otros ectotermos. La exposición a un frío severo y prolongado probablemente acabaría con la mayor parte de los endotermos vivientes, desnudos y tropicales, como los rinos, hipos, elefantes y armadillos, y también es muy posible que eliminara a los dinosaurios, los desnudos endotermos tropicales del mesozoico».
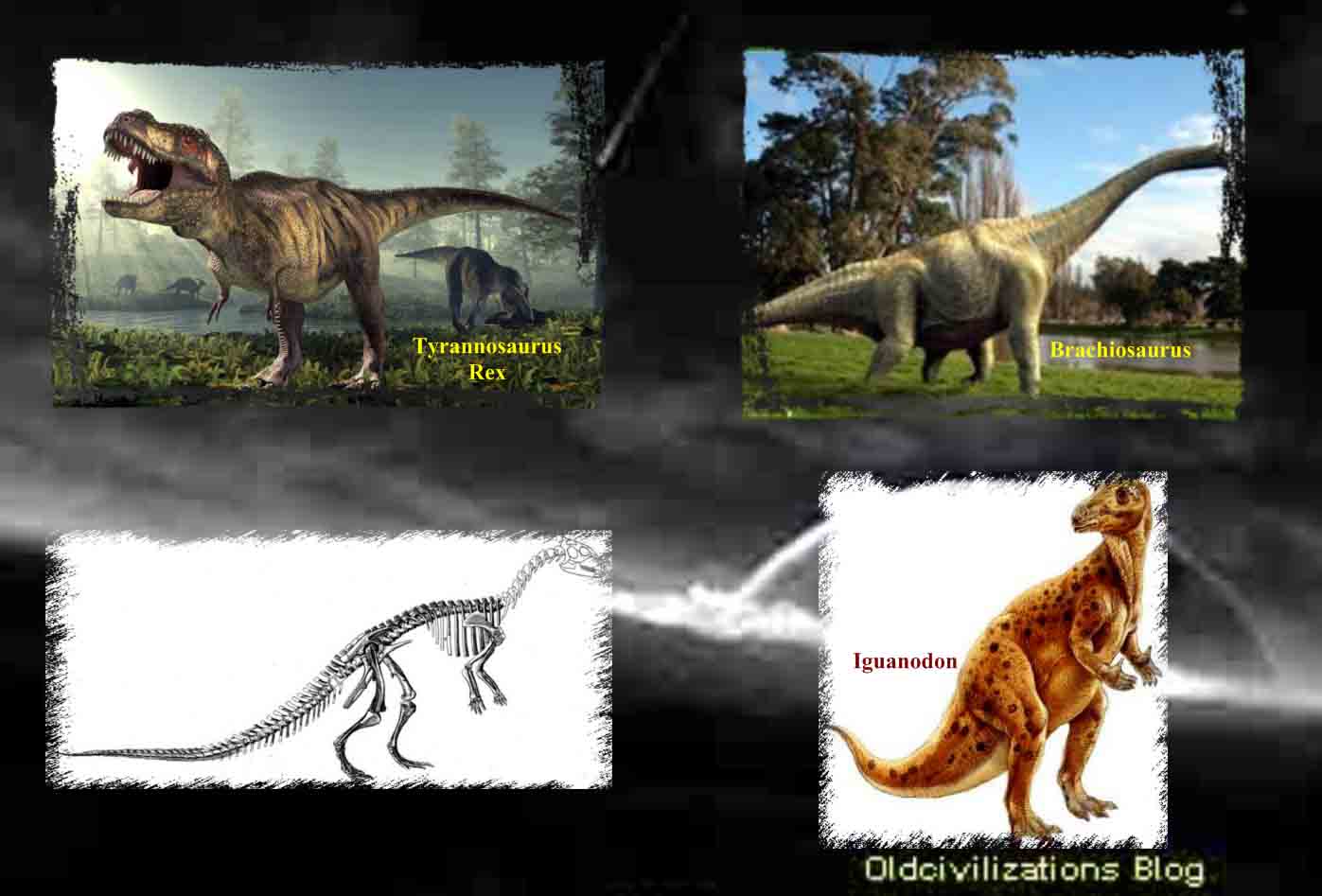
Por el contrario, dice Hotton, «si los dinosaurios hubiesen sido endotérmicos, algunos de ellos habrían sobrevivido». Si algunos de los pequeños dinosaurios hubieran dispuesto de plumas como aislante contra el frío, sobre lo cual se ha especulado mucho, pero nunca ha sido definitivamente establecido, habrían sobrevivido «en una diversidad comparable a la de los mamíferos y las aves supervivientes». Incluso algunos especímenes de los dinosaurios grandes y desnudos hubieran sobrevivido, aunque sólo fuera durante un corto tiempo. Sin embargo ningún dinosaurio supuestamente sobrevivió a la extinción masiva que tuvo lugar a fines del período cretáceo. Hotton concluye: «La supervivencia de los ectotermos, al igual que la de los endotermos, durante la transición cretáceo-terciaria, refuerza aún más el punto de vista de que existía una gran diferencia en la fisiología termal entre los dinosaurios y los tetrápodos vivientes». Aunque los argumentos en favor del gran tamaño y la homeotermia inercial como características controladoras de la fisiología del dinosaurio, parecen satisfacer a muchos paleontólogos, Bakker no emprendió la retirada. Los dinosaurios más pequeños se contaban entre los más ágiles y vitales de la especie, como ocurre en los endotermos. El primer grupo de tetrápodos que daba pruebas de elevada producción de calor, los terápsidos, pronto dieron auge a los grandes animales terrestres dominantes. Los dinosaurios parecen representar un elevado nivel de endotermia y reemplazaron a las dinastías de terápsidostecodontos. La entera historia de los fósiles de los grandes vertebrados muestra la naturaleza progresiva de la evolución entre los grandes tetrápodos. Los dinosaurios parecen haber alcanzado un nivel de rendimiento en la regulación térmica y en la locomoción comparable a la de muchos de los mamíferos del cenozoico posterior. Algunos de los pequeños dinosaurios depredadores parecen haber llegado a desarrollar cerebros del tamaño y la complejidad semejantes a los de las modernas aves no voladoras del mismo peso. Así reinterpretado, el éxito de los dinosaurios se convierte en parte de una progresión irreversible que conduce desde los primeros tetrápodos terrestres del devónico a la gran complejidad de los mamíferos modernos. Pareció establecerse el consenso en que no existía una simple estrategia termorreguladora común a todos los dinosaurios. Los grandes dinosaurios, los saurópodos, como el Brontosaurus, probablemente estaban más cerca de la ectotermia. Sus huesos sugerían que no fueron animales muy activos o ágiles y resulta difícil imaginar cómo los saurópodos podrían haber encontrado alimento suficiente para mantener un metabolismo elevado. La mayor parte de los dinosaurios, debido a su gran tamaño y al clima suave, probablemente mantuvieron temperaturas corporales bastante constantes. Eran homeotérmicos inerciales. Los pequeños dinosaurios, cuyos huesos sugieren una buena capacidad para la velocidad y la agilidad, pudieron haber sido endotérmicos reales. Desde luego los más pequeños como el Compsognathus, que fue un contemporáneo delArchaeopteryx, eran demasiado pequeños para poder ser homeotérmicos inerciales.
La maravilla de los dinosaurios es que son un enigma que parece por encima de toda solución. La ciencia ha identificado cientos de especies de dinosaurios, ha ensamblado sus huesos y encontrado la fecha en el tiempo de su presencia en la Tierra, pero son muchas las cosas relacionadas con las vidas de esas criaturas extrañas y monstruosas que desafían toda explicación. Los científicos salieron del debate sobre la sangre caliente al menos con un nuevo concepto del enigma del dinosaurio. Bakker, aunque fracasó en su empeño de probar que los dinosaurios frieron endotérmicos, inspiró a un abundante grupo de paleontólogos a la exploración de la ecología y la estructura comunitaria de esos reptiles tan poco corrientes y a tratar de comprender su lugar en la evolución de la vida. Incluso Nicholas Hotton, el más duro de los adversarios de Bakker en aquellos debates, tuvo que conceder: «Las estrategias termales alternativas y el estilo de vida al alcance de los dinosaurios pueden muy bien haber sido tan exóticos como las formas de sus cuerpos, de las que no hemos visto nunca nada comparable». Ostrom creía firmemente que las aves son descendientes directos de los dinosaurios. Su estudio del Deinonychus lo llevó a un análisis de docenas de otros dinosaurios, en particular de los pequeños carnívoros corredores conocidos como terópodos coelurosaurios. Las similitudes existentes entre el Archaeopteryx y los coelurosaurios fueron a su juicio sorprendentes. Con la excepción de las plumas el Archaeopteryx se parece más a un pequeño dinosaurio que a un ave moderna. Por lo tanto, concluyó Ostrom, debía haber una estrecha relación entre dinosaurios y aves, quizá más de lo que nadie se había atrevido a imaginar. Ostrom estaba resucitando la propuesta original de Thomas Henry Huxley de que existía un lazo entre los dinosaurios y las aves. Los científicos pudieron llegar a un consenso: las aves descendían de los reptiles. Durante muchos años del siglo XX, se abandonó la idea de Huxley de un ascendiente común. Los dinosaurios y las aves estaban remotamente emparentadas debido a que ambas especies descendían de un tronco común muy distante, identificado usualmente como los primitivos tecodontos. Éstos serían los ascendientes que a principios del triásico dieron lugar a todos los arcosaurios. Las aves, pues, no estaban más relacionadas con los dinosaurios que los cocodrilos lo están con los mismos dinosaurios o con los pterosaurios. En el caso de evolución paralela, dos organismos surgidos de un ascendiente común, divergen en dos ramas distintas que desarrollan semejanzas estructurales para adquirir capacidades comunes, como por ejemplo el vuelo. Las aves y los pterosaurios son ejemplos de evolución paralela. En el caso de evolución convergente, los animales de dos ramas independientes y sin ningún antepasado común, tales como murciélagos y aves, desarrollan similitudes al adaptarse al mismo tipo de nicho ecológicos, como el aire. Dado que algunos de los pequeños dinosaurios carnívoros y las protoaves compartían nichos ecológicos semejantes y perseguían el mismo tipo de alimento, es comprensible que desarrollaran anatomías similares. Pero de acuerdo con esa hipótesis eso no debía ser tomado necesariamente como signo de próximo parentesco.
Gerhard Heilmann, un paleontólogo danés, fue responsable de la extendida negativa a aceptar la existencia de un lazo directo entre el dinosaurio y el ave. En su libro más importante, The Origin of Birds, publicado en 1926, Heilmann reconoció «los sorprendentes puntos de semejanza» entre los coelurosaurios y las aves. Destacó uno de los muchos rasgos semejantes a los de las aves que se daban en el coelurosaurio: «Huesos huecos de estructura muy ligera, miembros traseros excesivamente largos con metatarsales fuertemente prolongados y un “dedo trasero”, una mano larga y estrecha, una cola larga y un cuello largo, órbitas largas y costillas ventrales». Pero faltaba uno de los rasgos más característicos de las aves. Observó que el coelurosaurio estaba falto de toda evidencia de los huesos del cuello (clavículas). Si elcoelurosaurio no tenía clavículas ¿cómo podían sus supuestos descendientes avícolas haberlo desarrollado? La ausencia de clavículas en el coelurosaurio, decidió Heilmann, «sería en sí misma prueba suficiente de que esos saurios no pueden ser los antepasados de las aves». En su lugar sugirió que los antepasados de las aves podrían encontrarse entre los pseudosuchianos, un grupo de tecodontos de principios del triásico. Ya antes, en 1913, Robert Broom había encontrado en África del Sur un pequeño pseudosuchiano bípedo que tenía una antigüedad de doscientos treinta millones de años, el Euparkeria, que parecía contar con todas las cualificaciones anatómicas necesarias para ser el antepasado de las aves. Primero en 1973 y más tarde en 1975, Ostrom se incorporó al debate con firmes argumentos en favor de la existencia de un lazo directo entre los dinosaurios y las aves. En parte lo hizo en respuesta a otra propuesta de un antepasado para las aves que negaba todo parentesco con los dinosaurios. Alick D. Walker, de la Universidad de Newcastle, sugirió que cocodrilos y aves, incluyendo elArchaeopteryx, procedían de un antepasado común teocodonto, pero que no era el mismo que daría lugar a los dinosaurios. Basaba esta idea en el parecido entre la caja cerebral y el cráneo de los cocodrilos actuales y las aves, así como algunos notables rasgos propios de las aves que se daban en el Sphenosuchus, una criatura de finales del triásico, hace doscientos veinte millones de años, que se pensaba era un cocodrilo primitivo o un tecodonto en el camino evolutivo hacia su transformación en cocodrilo. Ostrom pudo mostrarse más concluyenter de lo que lo había sido Huxley debido a que para entonces ya se habían descubierto y examinado muchos más dinosaurios pequeños, bípedos. Casi invariablemente sus extremidades, pies y garras, eran casi iguales a las del Archaeopteryx. Unos cuantos de loscoelurosaurios recién descubiertos parecían poseer el desaparecido ingrediente que tanto pareció importarle a Heilmann y, por lo visto, tenían clavículas. Una expedición polaco-mongol acababa de regresar con la prueba de que el Velociraptor, encontrado en el desierto de Gobi, parecía tener un par de huesos del cuello o clavículas. Lo mismo ocurría con el Segisaurus, un fósil norteamericano estudiado por Ostrom.

Al resumir las pruebas en favor de la ascendencia dinosáurica de las aves, Ostrom escribió en 1975: «Es más probable que el Archaeopteryx adquiriera su gran número de caracteres derivados de los terópodos, por convergencia o en paralelo al mismo tiempo que esos mismos caracteres estaban siendo adquiridos por algunos terópodos coelurosaurios, presumiblemente de un común antepasado. ¿O era más factible que esos muchos rasgos derivados fueran comunes a algunos terópodos pequeños y al Archaeopteryx debido a que este último procedía directamente de un terópodo semejante? No existe en absoluto ningún reparo en mi mente para aceptar que la última de las explicaciones es, con mucho, la más probable». La nueva actitud de Ostrom sobre el Archaeopteryx provocó un nuevo asalto en el debate de otra cuestión que hacía tiempo que dividía a los científicos. Se refería a uno de los mayores misterios de la evolución: el origen del vuelo en los animales. Como adaptación, el vuelo se coloca al mismo nivel que la salida del pez del mar, hace más de trescientos cincuenta millones de años, para convertirse en el primero de los vertebrados terrestres. El registro de los fósiles nos muestra que el primer vuelo real se desarrolló independientemente en varias ocasiones. Los primeros animales voladores fueron los insectos alados, que aparecieron hace unos trescientos cincuenta millones de años. Después fueron los pterosaurios que, probablemente, eran capaces de efectuar un vuelo motriz. Después llegaron las aves voladoras, quizá en una época tan antigua como la del Archaeopteryx, pero ciertamente no mucho después en el mesozoico. Los únicos otros animales voladores de propulsión propia, los murciélagos, cruzaron el aire por vez primera hace sólo cincuenta millones de años. Aquellos que se han sentido interesados y sorprendidos por el origen del vuelo han fijado su atención en las aves, las más llamativas y logradas de todas las criaturas aéreas. El descubrimiento del Archaeopteryx facilitó a los científicos el más claro y reconocido ejemplo de una criatura próxima o en plena transición en relación a la capacidad de vuelo. En 1880, Marsh sugirió que algunos reptiles treparon a los árboles para situar en ellos sus nidos, para alimentarse y para escapar de los depredadores. Esos primitivos antepasados de las aves tenían necesidad de saltar de una rama a otra y después de un árbol a otro. Consecuentemente cualquier variación anatómica que aumentara ese tipo de movilidad pasó a las subsiguientes generaciones por las leyes de la selección natural. De ese modo, presumiblemente los reptiles desarrollaron plumas rudimentarias en sus antebrazos para descender cuando saltaban de rama en rama y para caer al suelo como con la ayuda de un paracaídas. Al señalar ese punto Marsh citó el modelo de los modernos animales planeadores, como por ejemplo la llamada ardilla voladora. Más tarde, de acuerdo con la hipótesis de Marsh, estos reptiles protoavícolas, pudieron haber adaptado esas alas y fortalecido sus músculos para conseguir un vuelo impulsado muscularmente. Otros investigadores consideran que durante el gran espacio de tiempo abarcado por el triásico, del cual apenas si tenemos registros eficaces, debió de producirse un alargamiento gradual de los dedos externos y un gran desarrollo de las capacidades trepadoras, con lo cual se ayudaba al animal a correr más. El siguiente cambio hacia las plumas pudo haber sido sencillo. Las alas debieron de ser, al principio, utilizadas para correr, después para saltar y descender desde las alturas y, finalmente, para remontarse.
Franz von Nopcsa argumentó que las aves descendían de criaturas bípedas semejantes a los dinosaurios, que corrían en el suelo. Creía que para incrementar su velocidad de carrera aquellas protoaves desarrollaron extremidades anteriores con plumas, que podían mover hacia adelante y hacia atrás como remos. Probablemente esa habilidad no sólo permitió a aquellos animales correr con mayor rapidez sino que les dio la fuerza suficiente para elevarse y planear sobre distancias cortas. Nopcsa imaginaba que éste pudo ser el origen del vuelo de las aves. Pero eso resultaba aerodinámicamente imposible, como señaló un crítico. Tan pronto como el animal despegara del suelo sus patas no podrían seguir propulsándolo hacia adelante y caería de inmediato. Los científicos parecían sentirse más cómodos con la teoría arbórea. Podían imaginarse con mayor facilidad que los animales hubieran tratado de utilizar la gravedad para descender de las ramas que verlos luchando contra esta fuerza para elevarse del suelo. Tuvo que resultar importante, razonaron, el hecho de que todos los actuales vertebrados que vuelan son arbóreos. En sus estudios sobre el Archaeopteryx, Ostrom fue más allá de limitarse a estudiar su parentesco con los dinosaurios para presentar la cuestión del vuelo de las aves. Puesto que el Archaeopteryx carecía de esternón, el hueso pectoral que liga los músculos del vuelo, sintió dudas razonables sobre su capacidad, como ave primitiva, para desarrollar un vuelo monitorizado. Creía, más bien, que se trataba de un animal «ligero de pies, rápido y que probablemente habitaba en tierra y no en los árboles, como ocurre actualmente con los faisanes, las perdices o las codornices, por ejemplo, con cuyas garras las suyas tenían un extraordinario parecido». Todo eso le sugirió a Ostrom una nueva idea y especuló con que elArchaeopteryx y otras protoaves hubiesen sido depredadores que cazaban levantando sus presas, generalmente insectos, golpeándolas en el aire al cortarles el camino. Sus antebrazos emplumados, propuso, debían ser utilizados para golpear a los insectos en vuelo. Cuando la superficie emplumada de sus alas se desarrolló hasta alcanzar un mayor tamaño, esto les pudo conferir la capacidad de elevarse en el aire durante sus actividades depredadoras y, con el transcurso del tiempo, hizo posible que aquellas criaturas evolucionaran en el aire con vuelo a impulso propio. Gerald Caple, un farmacéutico, encontró poco probable que la red para cazar insectos pudiera haber evolucionado hasta dar impulso elevatorio a las capacidades necesarias para el vuelo activo motor, mostraron cómo los corredores bípedos podían desarrollar la fuerza y la capacidad suficiente para volar. Afirmaron que tales animales, corriendo y abalanzándose sobre sus presas, podrían haber utilizado sus extremidades anteriores para guardar el equilibrio, como hacen los seres humanos cuando dan un gran salto de longitud.
Esos movimientos de control se parecen rudimentariamente al golpe de ala en el vuelo. Si ese golpe de las extremidades superiores se llevaba a cabo cuando se daba un momento aerodinámicamente apto, como por ejemplo extremidades superiores cubiertas de plumas, el aire se deslizaría por encima y por debajo del ala a diferentes ritmos, con lo cual crearía un poder ascensional y prolongaría el salto. Predijeron que el ritmo que llevaría al vuelo sería rápido. De acuerdo con sus hipótesis, el animal no necesitaba desarrollar músculos nuevos y diferentes para llevar a cabo el vuelo motor, en contraste con las necesidades de un planeador arbóreo. Bock razonó de este modo: «Puesto que existen menos depredadores en los árboles que en el suelo, es posible que las protoaves se mantuvieran en los árboles mucho tiempo en condiciones que favorecían el desarrollo de una eficaz superficie aislante y, eventualmente, un cuerpo completamente cubierto de plumas. Más aún: si las protoaves utilizaban los árboles para anidar hubiera resultado muy ventajoso para los adultos de la especie el calentar los huevos en vez de dejarlos abandonados completamente para ser incubados por el calor solar. Por eso las fuerzas de la selección natural en los árboles pudieron favorecer la evolución hacia la endotermia y el aislamiento mediante plumas». Esta teoría, aunque muy poco probable, podría explicar otro aspecto intrigante de los dinosaurios y las aves:¿puede la sangre caliente de las aves ser presentada como evidencia en favor de que sus antepasados, los dinosaurios, también eran animales de sangre caliente? Bock parece sugerir que los reptiles ya en camino de convertirse en aves desarrollaron su sangre caliente sólo en las etapas finales de su transición hacia las aves. Robert Bakker ya había llegado a convencerse de que la endotermia no sólo era un rasgo común entre los dinosaurios, probablemente antes de su transición a las aves, sino que era una prueba convincente de la existencia de un estrecho parentesco dinosaurio-ave y, recíprocamente, que la sangre caliente de las aves era prueba evidente de que los dinosaurios fueron, también, animales endotérmicos. Para Bakker el asignar a los dinosaurios una categoría superior resultaba algo lógico y justificado. Había entrado a saco en la paleontología con su declaración de la «superioridad» de los dinosaurios y ahora trataba de definir lo que en ellos había de especial en términos científicos. Al igual que los mamíferos, arguyó, los dinosaurios habían dado un salto evolutivo trascendental con respecto a sus antepasados. Los mamíferos se habían desarrollado a partir de los reptiles, progresando desde la ectotermia a la endotermia y lo mismo habían hecho los dinosaurios. Además no tenía más sentido separar a las aves de los dinosaurios del que tendría separar a los murciélagos de los mamíferos. Significa, como Ostrom escribiría en cierta ocasión, que los «dinosaurios no quedaron extinguidos sin dejar descendientes». Y que las plumas que crecieron en algunos dinosaurios no sólo llevaron al vuelo sino que fueron aislantes térmicos que a su vez fueron la razón primaria del éxito de los descendientes de los dinosaurios.
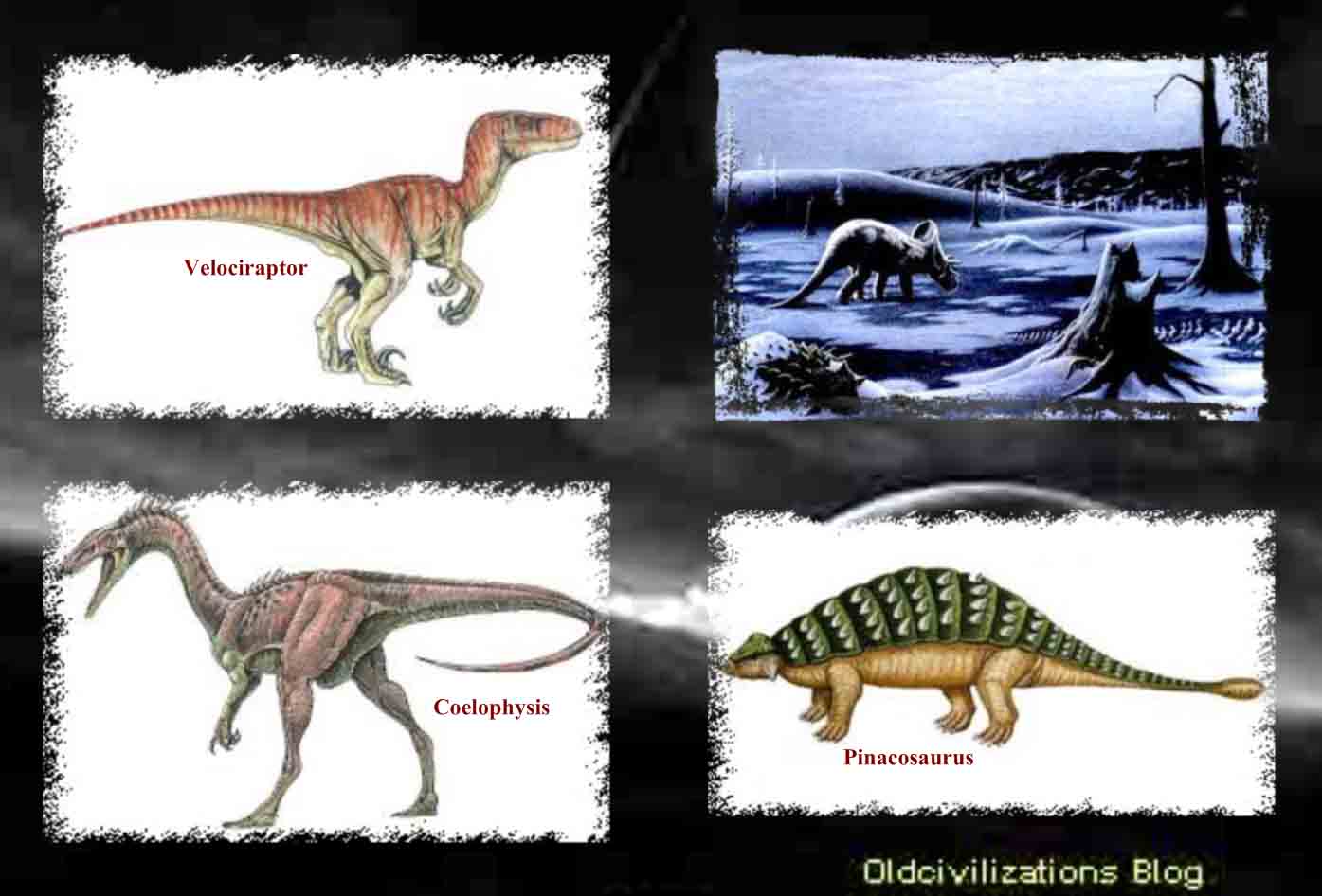
John R. Horner y Robert Makela, ambos paleontólogos, se dedicaron a cavar en piedra arenosa de color marrón grisáceo. Al cabo de pocos días, trabajando bajo un sol ardiente y un fuerte viento, excavaron los restos de un nido de barro que contenía quince pequeñas crías de dinosaurios fosilizadas. Cada una de las crías tenía aproximadamente un metro de longitud. Éstas fueron las primeras crías de dinosaurio encontradas en su nido y sus dientes mostraban que llevaban algún tiempo comiendo. Eso señaló el comienzo de uno de los descubrimientos de dinosaurios más brillantes del siglo XX. Horner y Makela descubrieron más de trescientos huevos de dinosaurio en distintos nidos. Algunos huevos conservaban el esqueleto del embrión. Los dos buscadores sacaron más de sesenta esqueletos, totales o parciales, incluyendo los de las crías, y otros de dinosaurios muy jóvenes, así como los de sus padres. Nada de lo que había sido desenterrado hasta entonces había aportado un material básico informativo para poder conocer la conducta social de los dinosaurios. Así se obtuvo la primera prueba evidente de que los dinosaurios se ocupaban del cuidado de sus crías, un rasgo característico que no se da en absoluto en los reptiles actuales. Más tarde, en el verano de su primera temporada de trabajo, Laurie Trexler, que los estaba ayudando, encontró allí la cabeza de un dinosaurio adulto. «Si me tengo que conformar con una parte de un animal -explicó Horner-, ciertamente prefiero que sea el cráneo». Aquél le intrigó aún más, pues parecía distinto de todos los dinosaurios conocidos hasta entonces. Durante meses Horner examinó el espécimen. Finalmente decidió que pertenecía a un miembro de la subfamilia de los arcosaurios conocida como hadrosauridae, el primero de los dinosaurios de pico de pato que fuera descubierto. Eran los más abundantes y diversos de los dinosaurios del cretáceo. La mayor parte de los otros hadrosaurios tenían una abertura de las fosas nasales muy grande que incluía las órbitas de los ojos. Aquél, por el contrario, tenía unas fosas nasales pequeñas con un hueso muy grande entre éstas y las órbitas. A ese respecto, el animal se parecía al Iguanodon, pero éste tenía dientes pequeños, sencillos y escasos. El cráneo que intrigaba a Horner tenía una verdadera batería de cientos de grandes dientes unidos entre sí y que eran mucho más adecuados para una mejor masticación y el roído de la vegetación dura. Esta característica y algunas otras sugerían que el animal debió de tener entre los ocho y los diez metros de longitud. Finalmente llegó a la conclusión de que se trataba de un hadrosaurio de un nuevo género y especie, a los que bautizó con el nombre de Maiasaura peeblesorum. Maiasaura significa algo así como «la buena madre lagarto», nombre que parece muy apropiado a partir de la evidencia.
Horner y Makela continuaron buscando y excavando juntos en años sucesivos. De este modo pronto hicieron descubrimientos suficientes para darse cuenta de que no habían descubierto un nido aislado, o dos, sino toda una colonia. Pronto excavaron dos nidos más con crías dehadrosaurios. En uno de los nidos las crías eran aún más jóvenes que los encontrados anteriormente. Medían menos de medio metro. Otras eran de más edad y habían crecido hasta los dos metros de longitud, lo que daba a Horner nuevos argumentos para creer que aquellas crías y pequeños eran cuidados y atendidos por sus padres durante varios meses después de la incubación y la ruptura del huevo. El Maiasaura, «la buena madre lagarto», debió de haber alimentado a sus hijos, regresando frecuentemente al nido de fango arenoso con semillas, bayas y hojas verdes. El equipo de Horner también encontró los restos destrozados por el tiempo de nueve nidos vacíos, todos los cuales contenían gran abundancia de trozos de cáscaras de huevos fosilizadas. Aquellos huevos de hadrosaurio, una vez fueron reconstruidos, medían unos veinte centímetros de longitud con la forma de elipsoides chatos, aproximadamente del tamaño y la forma de los huevos del Protoceratops que anteriormente habían sido hallados en el desierto de Gobi. Las cáscaras eran algo más gruesas que las de un huevo de gallina, más bien como las de un huevo de avestruz. Las superficies externas de las cáscaras eran rugosas. Los nidos estaban separados entre sí unos siete metros como mínimo, lo que Horner observó con especial atención, porque siete metros suele ser, aproximadamente, la longitud del hadrosaurio adulto. Todos los nidos ocupaban la misma capa o estrato, lo que parecía indicar un mismo «horizonte temporal». Eso hacía suponer que aquel lugar había sido sede de una colonia de crianza semejante a las de algunas aves modernas. En 1979, mientras cavaba en un montículo próximo, Horner descubrió otra colonia de nidos. Por esa razón el montículo fue bautizado con el nombre de Egg Mountain (montaña de los huevos). Los diez nidos encontrados allí contenían los restos de nada menos que veinticuatro huevos cada uno. Aquellos huevos, con superficies exteriores lisas, eran algo más pequeños y más elipsoidales que los hallados anteriormente. Estaban colocados de modo circular con el extremo más puntiagudo enterrado en el fondo arenoso del nido. Terminado el empollamiento la criatura salía por la parte superior del huevo, dejando tras sí la parte baja intacta entre los sedimentos. A partir de los huevos y de algunos huesos, Horner llegó a la conclusión que aquella colonia había sido ocupada por otra especie de dinosaurio, probablemente una especie del algo más pequeñoHypsilophodon. Horner estuvo en condiciones de sacar dos deducciones a partir de la colonia de nidos de la Egg Mountain. Los nidos se encontraban a tres niveles diferentes sobre el suelo. Esto le sugería la idea de que los adultos habían ido anidando en el mismo lugar, año tras año, durante bastante tiempo. Es decir, era el lugar al que regresaban en cada temporada de crianza.
Al advertir la ausencia de restos de jóvenes en aquellos nidos, así como el hecho de que las partes inferiores de los huevos estaban bien conservadas y no rotas como en los nidos del anterior descubrimiento, sin duda a causa del pataleo y movimiento de los pequeños, Horner dedujo que los jóvenes de esta especie no se quedaban en el nido tanto tiempo como los de la otra y lo abandonaban pronto tras haber salido del huevo. No obstante había signos que indicaban la atención de los padres. Restos de entre quince y veinte esqueletos pequeños, de longitudes que iban desde medio metro a un metro y medio fueron encontrados unos cerca de otros. Si las crías abandonaban el nido inmediatamente después de salir del cascarón, es posible que por casualidad algunos hubieran muerto allí, pero un grupo de quince o veinte parecía poco probable. Esa circunstancia recordó a Horner los lugares próximos al nido en los que se desarrollan algunas aves de nuestro tiempo. Como informó en Nature, aquellos jóvenes dinosaurios «o bien seguían en la colonia o regresaban al lugar frecuentemente». Ambas cosas hubieran sido indicios de la existencia del cuidado paternal. Los análisis del desarrollo de los huesos de las crías de dinosaurios llevaron a Horner a nuevas evidencias. Descubrió la existencia de un crecimiento rápido. El metabolismo de aquellas criaturas parecía haber tenido el ritmo rápido de los animales de sangre caliente más que el lento y perezoso de los reptiles de sangre fría. Su razonamiento era que los nidos de los hadrosauríos, en particular, contenían crías en los cascarones que ni siquiera llegaban a medir medio metro, así como restos de animales muy jóvenes de un metro y fragmentos de otros que podrían haber llegado a los dos metros. O bien crecían muy rápidamente o seguían mucho tiempo en el nido. El cocodrilo joven, que es un animal de sangre fría, así como el arcosaurío, crecen un tercio de metro por año. Incluso las especies de más rápido crecimiento entre los animales de sangre fría tardarían un año en conseguir el tamaño de los jóvenes hadrosauríos descubiertos por Horner. Parece improbable que los jóvenes dinosaurios se quedaran en el nido tres años, o incluso uno, por muy cariñosos y dispuestos a alimentarlos que estuvieran sus padres. Más bien habría que pensar que los dinosaurios crecían a un ritmo comparable al de los avestruces, por ejemplo, que es la criatura de sangre caliente de más lento crecimiento. Un avestruz alcanza un tamaño de un metro en un período de entre seis y ocho meses. Por alguna razón las crías de la buena madre lagarto de Montana parecían capaces de crecer a ese ritmo o incluso a otro más rápido y esto podría indicar que se trataba de animales de sangre caliente.

Horner dijo que la gente había buscado en lugares equivocados, ya que la mayor parte de los paleontólogos se habían encerrado en la idea de que los dinosaurios eran criaturas de las cuencas de los ríos bajos, llanuras costeras y tierras pantanosas y por lo tanto fue en esos lugares en donde buscaban todo lo que pudiera relacionarse con los dinosaurios. Casi todos sus hallazgos de huesos habían sido hechos en tales sedimentos. Sin embargo, Horner encontró sus nidos en terrenos que fueron más elevados y más áridos. De acuerdo con la hipótesis de Horner los dinosaurios emigraron de las tierras bajas, situadas a unos cien kilómetros, para hacer sus nidos, poner sus huevos y cuidar a sus crías en lugar alejado y seguro, que protegiera a sus hijos de los dinosaurios carnívoros. Los primeros aspectos de la forma de vida de los dinosaurios que pudieron ser deducidos con cierto grado de certeza, como hemos podido ver, se relacionan con la locomoción y la manutención. Si las extremidades delanteras del animal habían sido considerablemente más pequeñas que sus miembros traseros, podía presumirse que esto los hacía bípedos. Esto podía ser cierto si los miembros anteriores terminaban en manos con garras afiladas para agarrar y atacar. Pero algunos dinosaurios tenían unos crecimientos óseos, parecidos a cascos, en los dedos de los miembros más cortos y, por lo tanto, eran cuadrúpedos, o al menos lo fueron durante algún tiempo. Los grandes saurópodos, con cuatro miembros fuertes, como columnas y patas de elefante, eran indudablemente cuadrúpedos. Los rastros de sus huellas fosilizadas lo confirman. La forma de los huesos de las extremidades y la configuración de las caderas y los hombros, por otra parte, prueban que los dinosaurios se alzaban más rectos y erguidos que otros reptiles que mantienen su postura clásica de animales reptantes. Los dientes también nos ofrecen claras indicaciones que nos ayudan a establecer su dieta. Si los animales tenían pequeños dientes más bien planos esto indicaba que se alimentaban de vegetales blandos. Si los dientes eran mayores y estaban situados en las mandíbulas para trabajar en la masticación como si fueran tijeras, los animales podrían haber comido vegetación más dura y fibrosa que requería ser cortada y triturada. Algunos dinosaurios no tenían en absoluto dientes, lo que indica que subsistían alimentándose de insectos, huevos y frutas muy blandas. Aquellos otros con los dientes más amenazadores, largos, agudos y en forma de sierra, eran carnívoros que cazaban a los herbívoros mientras éstos pastaban, lo mismo que los leones cazan en la actualidad. Los huesos delatan también los gustos alimenticios de los dinosaurios. Los huesos más densos y laminados pertenecen a los carnívoros. En algunos pocos casos los científicos han encontrado pruebas directas de la dieta de un dinosaurio. Fue hallado un anatosaurio momificado que murió con el estómago lleno y su contenido se componía de agujas de coniferas, ramas, frutos y semillas.
Es posible que algunos dinosaurios ingirieran rocas, como algunas aves tragan finos guijarros para ayudarse en su digestión. Los cazadores de esqueletos han encontrado frecuentemente piedras extremadamente suaves, mezcladas con los esqueletos de los saurópodos. Se las llama gastrolitos (piedras gástricas). Lo más probable es que esas piedras fueran tan finamente pulidas por las aguas de algún riachuelo existente en otros tiempos y no por los jugos gástricos de los dinosaurios. A lo largo de los años los científicos han intentado obtener cierta idea sobre la fisiología y la conducta del dinosaurio basándose en el tamaño y la forma de sus cerebros. Sus cerebros eran extremadamente pequeños en relación con sus cuerpos y consecuentemente, su inteligencia no debió de ser muy alta. Pero algunos de sus sentidos podrían compensar algo de esta deficiencia mental. Un cuidadoso examen de la estructura cerebral ha revelado la existencia de bulbos olfativos y lóbulos ópticos bien desarrollados, lo que indica una buena capacidad olfativa y de visión. La estructura ósea interna de las orejas sugiere un excelente oído. Posiblemente estaban capacitados para oír notas extremadamente altas, quizá los agudos grítitos de sus crías, una habilidad muy importante para mantener la cohesión familiar. Todos los dinosaurios tienen órbitas demasiado grandes para sus ojos. ¿De qué color? Nadie lo sabe, pero los actuales reptiles tienen ojos de un color que va del rojo al amarillo. No hay ninguna prueba de que los dinosaurios pudieran expresar oralmente los pensamientos que pudieran cruzar por sus pequeños cerebros. Pero sí hay presunciones en ese sentido. Algunos huesos de sus cráneos sugieren que tenían una voz potente. W. E. Swinton, un paleontólogo británico, ha escrito: «En este aspecto podrían haber igualado a los cocodrilos, todos los cuales tienen un corto y agudo croar, o una especie de ladrido, mediante el cual se identifican entre sí en la oscuridad o que utilizan para indicar cuándo están irritados». Los dinosaurios, como todos los animales, debían de tener sus medios de ataque, de defensa o de huida ante los ataques. Los herbívoros que pastaban en las llanuras tenían que confiar en su vista y en su oído para descubrir a los depredadores y poder escapar, al tiempo que croaban, ladraban o dejaban escapar su sonido de tuba, para alertar a los otros del inminente peligro. Otros contaban para defenderse con sus garras, sus aguzados espolones, sus colas que utilizaban como trancas y sus cuernos aguzados, útiles tanto para el ataque como para la defensa. Los Triceratopsposiblemente luchaban entre sí como los cameros, empujándose y golpeándose con sus cráneos y sus fuertes cuernos. Sus esqueletos a veces mostraban señales de profundas heridas en la coraza ósea que se proyectaba hacia atrás como un escudo sobre el cuello y los hombros.
En 1971 la expedición polaco-mongola encontró en Gobi los restos de dos dinosaurios que, posiblemente, se habían matado entre sí. El depredador, un pequeño y rápido Velociraptor; estaba abrazado sobre su presa, un Protoceratops acorazado. El Velociraptor tenía un espolón en forma de hoz y muy afilado en cada una de sus patas traseras, que se parecía mucho al delDeinonychus de Ostrom. Uno de los espolones del depredador estaba hundido en la región que debió de ser el vientre de la presa. No estaba claro cómo el Protoceratops había logrado su póstuma venganza. Sin embargo este descubrimiento fue considerado como una prueba más de la nueva teoría que consideraba a los dinosaurios como más ágiles y activos que los reptiles ordinarios. Más aún se ha podido deducir de las huellas dejadas por los dinosaurios. Las huellas de las patas, como en cierta ocasión diría Richard Swann, «son fósiles de seres vivos, mientras que todos los demás fósiles son reliquias de los muertos». De las series de huellas de patas los científicos han deducido la velocidad de los dinosaurios corredores, han comprobado cómo los depredadores perseguían a sus presas, los saurópodos, deduciendo datos que los han llevado a su primera comprensión de la conducta en grupo de los dinosaurios. Éstos, al parecer, viajaban frecuentemente en rebaños, por lo cual podrían ser animales gregarios. Uno de los descubrimientos más importantes debidos a las huellas rescata en cierto sentido al Brontosaurusdel agua, y lo sitúa en tierra firme. Algunos libros ilustrados aún representan a los Brontosauruscon sus hocicos en aguas pantanosas, en actitud letárgica, casi inmóvil, masticando plantas acuáticas blandas. Los paleontólogos parecían creer que éste era el papel reservado en la vida a los Brontosaurus. Sus patas no eran lo suficientemente robustas para servir de apoyo a sus pesados cuerpos, alegaban los científicos, y por lo tanto necesitaban la flotabilidad de sus cuerpos dentro del agua. Además estaba la abertura nasal en la parte superior de la cabeza, lo que al parecer permitía respirar al animal aun con su cuerpo completamente sumergido. Todo esto dio lugar a la imagen lenta y pesada del Brontosaurus presentándolo como una especie de hipopótamo de los dinosaurios, aunque aún más lento y perezoso. La imagen comenzó a cambiar -en la mente de los científicos después de que Roland T. Bird, del Museo Americano de Historia Natural, examinó algunas huellas de dinosaurios en Texas, en la década de los años 1940-1950. Bird dio con la secuencia de huellas del mismo modo que Horner encontró las crías de dinosaurios.
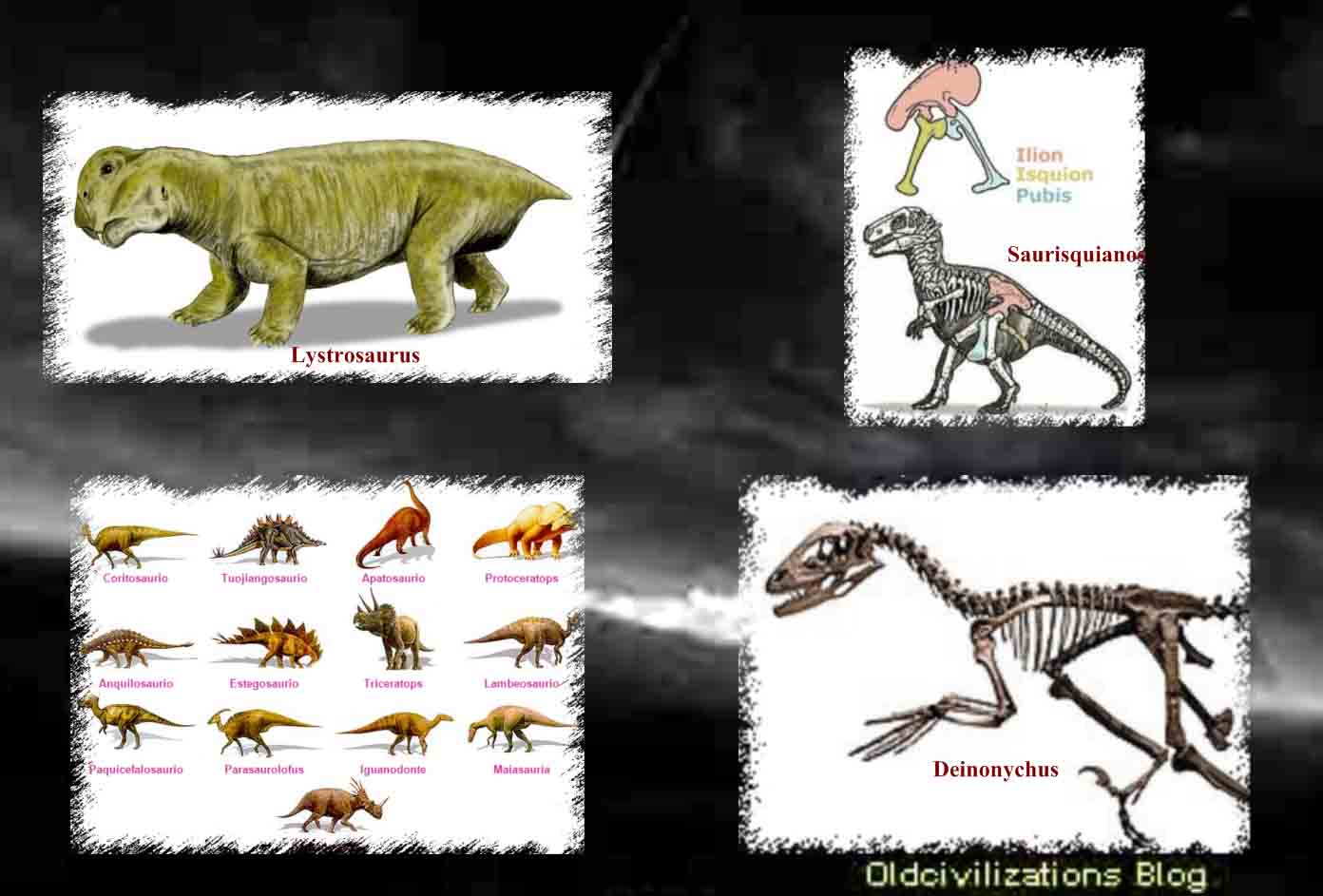
Bird encontró algunas huellas de patas de cuatro dedos, la primera serie de huellas de unBrontosaurus. Cada una de las huellas tenía un metro de longitud y estaban impresas con fuerza, profundamente, en lo que debió de ser una superficie fangosa parcial o totalmente expuesta al aire. Las huellas, separadas entre sí por dos metros, eran definitivamente las de un animal terrestre. El Brontosaurus podía andar sobre sus cuatro patas. En otro lugar de Texas, Bird vio una continua sucesión de huellas de las patas del Brontosaurus en la roca. El animal había arrastrado su pesada cola sobre la superficie fangosa, otra señal de que andaba y no nadaba o flotaba a medias en el agua. De estas pruebas los paleontólogos dedujeron algo que ya hacía mucho tiempo que debieron saber: que el Brontosaurus no era un animal prisionero del agua. Sus fósiles no fueron hallados, según pudo comprobarse, en sedimentos de origen pantanoso, sino en los mismos sedimentos procedentes de aluviones que contienen los restos de los demás dinosaurios que están clasificados como animales terrestres. El estudio de las huellas fósiles es una rama de la paleontología que se conoce con el nombre de ichnografia, derivado de la palabra griega que significa huella o rastro. Las primeras sendas marcadas por las huellas de los dinosaurios que llamaron la atención de modo más general fueron las impresiones de patas parecidas a las de las aves descubiertas en el valle del río Connecticut en Massachusetts, en 1802, cuando los dinosaurios aún eran desconocidos. Desde entonces han sido descubiertos cientos de otras series de huellas en Nueva Inglaterra, Texas, Brasil, Australia y, más recientemente, en el cañón del río Peace en la Columbia Británica. Naturalmente las huellas de los dinosaurios son más abundantes que los huesos de dinosaurio. Mediante un análisis realizado por los científicos en las huellas, utilizando una fórmula establecida por R. McNeill Alexander de la Universidad de Leed, determinaron que algunos dinosaurios eran capaces de moverse con bastante rapidez. Algunos carnívoros de tamaño medio marchaban a una media de 16,5 kilómetros por hora, lo cual se aproxima a la máxima velocidad de carrera conseguida por el ser humano. Algunos de los carnívoros más lentos lo hacían entre los 6 y los 8,5 kilómetros por hora. Los herbívoros incluso podían ser más lentos con velocidades máximas de 6 kilómetros por hora. Con estos cálculos un ex discípulo de Ostrom, James O. Farlow, especialista en dinosaurios independiente, de Michigan, informó en 1981 que algunos carnívoros habían dejado huellas en Texas que demostraban que alcanzaban una velocidad de 42 kilómetros por hora, es decir, más rápidos que los seres humanos, pero que no podían competir con los caballos de buena raza ni con los galgos. Otros cálculos indicaban que en caso necesario el Tyrannosaurus podía superar los 45 o 50 kilómetros por hora.
Según se deduce de las huellas, los Tyrannosaurus solían marchar solos o en parejas. Pero el instinto de rebaño o manada debió de ser más fuerte en otras especies. En el Parque Estatal de Dinosaurios de Connecticut, muchas de las huellas transcurren paralelas como si los dinosaurios marcharan en rebaño. En las sendas dejadas por los saurópodos en Texas y descubiertas por Roland Bird también existen pruebas de que marchaban en rebaño. Bakker en un reciente reexamen de esas huellas cree haber reconocido señales de un rebaño «estructurado», con los más jóvenes y las crías en el centro rodeados y protegidos por los adultos. En Montana, Jack Horner encontró pruebas de que sus hadrosauríos no sólo constituían colonias ponedoras sino también que continuaban constituidos en grupo durante la mayor parte, o la totalidad, de su vida. En varios lugares encontró restos de jóvenes de unos tres metros de longitud con restos de adultos de seis o siete metros. En un lugar había veinticuatro jóvenes junto a diez adultos. Es posible que no parezca demasiado importante el saber que algunos dinosaurios viajaban en manadas o en rebaños y que otros cuidaban a sus crías, pero lo cierto es que esa conducta los diferencia claramente de otros reptiles. La mayor parte de los reptiles, como los peces y los anfibios, abandonan sus huevos tan pronto como han sido puestos. Algunos peces y reptiles, entre ellos el cocodrilo, guardan sus huevos hasta que termina la incubación o un poco después y una vez nacidas las crías tienen que alimentarse y cuidarse por sí solas. Ninguno de los reptiles modernos vive en rebaños o en manadas de caza ni en ningún otro tipo de comunidad social. Los dinosaurios fueron diferentes. Algunos de ellos tenían vida familiar y un sentido de comunidad. Horner creía que la conducta social que podía inferirse de las huellas y los nidos podía ser de gran importancia en el éxito tremendo de los dinosaurios que hizo posible que su existencia se prolongara durante ciento sesenta millones de años. Los sociobiólogos creen que ciertos comportamientos fundamentales pasan de una generación a otra por medio de los genes. Uno de esos rasgos, el altruismo en el reparto de los alimentos, se observa de manera primordial en las aves y en los mamíferos. Si los padres delMaiasaura llevaban la comida a sus crías en el nido, como Horner creía, era posible que su especie y algunas otras especies de dinosaurios se transmitieran el altruismo de modo genético. Sin ese altruismo y cierto instinto social, en el cual se incluía el reparto de la comida, era más que posible que un buen número de dinosaurios no hubiera podido llevar una vida próspera en un mundo lleno de depredadores.
Los hadrosaurios, por ejemplo, eran herbívoros bípedos que no disponían de ningún medio de defensa obvio, ya que carecían de cuernos, de garras o de protección corporal. Tampoco eran capaces de correr a gran velocidad, puesto que eran caminantes lentos. Debieron de ser muy vulnerables ante el ataque de los depredadores, sobre todo cuando estaban solos. Consecuentemente, y como las nuevas pruebas parecían indicar, se vieron obligados a formar rebaños y a cuidar de sus crías durante varios meses después de que salieran de su cascarón, guardándolos en colonias relativamente bien protegidas. Y debieron hacerlo bien puesto que pese a su vulnerabilidad los hadrosaurios se cuentan entre los dinosaurios más abundantes que existieron. Estaba claro que sobrevivía el número suficiente de jóvenes hadrosaurios para reemplazar a los adultos y mantener la población. Según dijo Horner en 1983 esto fue así porque probablemente supieron emplear una «estrategia de supervivencia comparable a la más común entre las aves, la construcción de colonias de anidamiento». Ponían menos huevos que otros reptiles, pero protegían sus nidos durante la incubación y se dedicaban a alimentar a sus crías. La anidación en colonia y la protección de los nidos es practicada por los cocodrilos, pero éstos no alimentan a sus crías ni comparten los alimentos. Para los hadrosaurios, argüía Horner, ambas cosas se hacían necesarias pues de otro modo hubiera sido más que cuestionable la supervivencia de una o dos de esas pequeñas y pacíficas criaturas de cada camada de veinte o veinticinco. El hecho de que en sus grupos o rebaños hubiera al menos dos jóvenes por cada adulto sugiere una supervivencia del ocho por ciento de cada camada. El hecho de que en los nidos se encontrara un número tan grande de crías sugiere que eran los padres quienes se cuidaban de alimentarlos. El salir de sus nidos para alimentarse por su cuenta habría significado para las crías el riesgo de ser devoradas por los depredadores o ser pisoteadas por los adultos. Como muchas aves, los pequeños hadrosaurios permanecían en sus nidos por instinto. No sabemos durante cuánto tiempo, pero debió de ser durante varios meses, hasta alcanzar una longitud de metro y medio como mínimo. Aquellas quince crías encontradas por Horner en su primer nido debieron de quedarse en él mientras sus padres iban a buscarles comida. Sus padres no regresaron a causa de cualquier desastre, pero las crías se quedaron en el nido hasta que les sobrevino la muerte por inanición. Es posible que esa conducta de cuidado paternal y compartir los alimentos estuviera restringida a sólo unas pocas especies, concluye Horner, pero en todo caso refleja una amplia «divergencia sociobiológica» entre las dos grandes ramas de los dinosaurios, los omitichianos y los saurischianos.

Otra especie de la que se sabe que ponía sus huevos en nidos circulares con una notable atención, como vio Roy Chapman Andrews, era la de los Protoceratops, también omitichianos. Los huevos redondos de un dinosaurio parecido al Brontosaurus, quizá un Hypselosaurus, eran puestos en largas filas lineales que no es la mejor forma si se piensa en alimentar a las crías en el nido. Podría ser, según dijo Horner, que los omitichianos pusieran sus huevos en círculo y lossaurischianos en filas. La conducta en cierto modo más sofisticada y social de algunos dinosaurios puede haber sido una adaptación evolucionaría relacionada con el herbivorismo. Aunque es posible que unos pocos carnívoros cazaran en manada, los dinosaurios que según se sabe se desplazaban formando rebaños eran saurópodos herbívoros. «Incluso si se trataba sólo de reuniones y no de rebaños en el sentido estricto del término, se trata de un comportamiento que no vemos en otros reptiles -afirmó Horner-. Eso es algo que sólo conocemos en los mamíferos y en las aves». Y lo que es más, subraya Horner, los omitichianosfueron «el primer gran grupo de herbívoros que habitó la Tierra y para poder sobrevivir, competir y muy especialmente lograr un éxito tan considerable, tuvieron necesidad de establecer un orden social fuerte». Pero algo ocurrió hace sesenta y cinco millones de años, a finales del cretáceo, algo tan devastador que alteró el curso de la vida en la Tierra. De modo aparentemente repentino, en relación con el transcurrir del tiempo geológico, casi la mitad de los géneros vivientes desaparecieron. Ninguna de las especies que vivían exclusivamente en tierra y que pesaban más de veinticinco kilos parecen haber sobrevivido. Y los más notables de los desaparecidos fueron los dinosaurios. Aunque los fósiles de una variedad de dinosaurios han sido encontrados en las capas superiores de las rocas del cretáceo, ni uno solo ha sido desenterrado de las capas del terciario que están inmediatamente encima. Los dinosaurios habían desaparecido para no volver a ser vistos jamás. Las extinciones a finales del cretáceo hicieron caer el telón de la Era de los Reptiles. Pero muchos dinosaurios habían muerto con anterioridad. El Stegosaurus, de piel blindada, se extinguió antes de que comenzara el cretáceo. Los gigantescos Diplodocus, Brachiosaurus y Brontosaurus se extinguieron en el transcurso del cretáceo antes de que se produjera la gran agonía. Todos o casi todos lossaurópodos de ese tipo habían desaparecido. La mayor parte de los iguanodontes se había extinguido para entonces, pero los dinosaurios aún existían hacia finales del cretáceo. Entre otros arcosaurios, los cocodrilos marinos y muchos ictiosaurios posiblemente se habían extinguido ya, pero los plesiosáuridos, los pterosaurios y los mosasáuridos parecen haber tenido su período de florecimiento a finales del cretáceo y después se extinguieron. Sólo algunos crocodílidos, entre todos los arcosaurios, se las arreglaron para cruzar esa frontera tan plena en acontecimientos que separa el cretáceo del terciario y han sobrevivido hasta nuestros días.
Ese algo que ocurrió hace sesenta y cinco millones de años es uno de los misterios más intrigantes de la ciencia. No obstante, los dinosaurios tuvieron una larga y próspera estancia en la Tierra antes de desaparecer. Los intentos más serios de resolver el misterio se enfocan en las condiciones generales. No fueron los dinosaurios en sí los que fallaron sino que se trató de algo de mayor importancia. Especies de animales que habían sobrevivido durante millones de años tienen que haberse adaptado bien al medio ambiente, pues de otro modo no hubiesen sobrevivido sin que cambiaran las condiciones de su existencia. Por lo general, los dinosaurios fueron criaturas con gran éxito. Debían continuar sobreviviendo en tanto que no cambiaran las condiciones de su existencia. Si se modificaban lentamente, sus organismos podrían no tener tiempo de adaptarse a esos cambios, mediante el normal proceso de la evolución. La extinción se produce a causa de cambios, de cambios repentinos, un tipo de cambios cuyos efectos no pueden ser asimilados por los organismos individuales en el transcurso de pocas generaciones. Cuando Walter Álvarez encontró la pista, estaba buscando otra cosa. Fue uno de esos descubrimientos casuales que salpican los anales de la ciencia. A partir de 1973, Álvarez y algunos otros colegas acudían cada verano a los Apeninos, en la región de Umbría en el centro del norte de Italia, para estudiar las extraordinarias formaciones de piedra caliza que allí se alzan. La roca está formada por sedimentos de las profundidades del mar acumulados durante un período de ciento cincuenta y cinco millones de años, desde el jurásico hasta el oligoceno, hace treinta millones de años. El mar cedió paso a la tierra firme y hace unos pocos millones de años la roca que se formó en el fondo marino fue alzada en un fenómeno de elevación de montañas para quedar expuesta a la erosión durante millones de años. La naturaleza muestra allí una de sus más bellas creaciones, profundos lechos de piedra caliza rosada que brindaban placer estético mucho antes de que llegara la revelación científica. En la garganta Bottaccione, cerca de la ciudad medieval de Gubbio, esa roca caliza destaca de modo particular y es fácilmente accesible. Y fue allí donde Álvarez aplicó su habilidad como geólogo para estudiar una cuestión relacionada con la historia de la Tierra. Los dinosaurios estaban entonces muy lejos de su pensamiento. Su interés era paleomagnético. Álvarez y sus colegas estaban buscando pruebas en las rocas que sirvieran como evidencia de una inversión en el campo magnético de la Tierra. Querían comparar las inversiones encontradas en Gubbio con las pruebas de otras inversiones que habían sido detectadas en la corteza de los fondos oceánicos por medio de magnetómetros operados desde los puentes de los buques. Partiendo de esos descubrimientos confiaba en poder desarrollar, tanto en tierra firme como en el mar, un calendario seguro y completo de las fluctuaciones magnéticas en el curso de la historia de la Tierra. Durante sus investigaciones, los geólogos trataban también de relacionar las inversiones de campos magnéticos con la evolución y la extinción de organismos, como mostraba el registro fósil. La piedra caliza de Gubbio es rica en foraminíferos, criaturas diminutas cuyas conchas de carbonato de calcio son la materia prima de una gran parte de la piedra caliza del mundo. Con una lupa manual los geólogos pudieron ver la profusión de microorganismos que se abrían camino a través de las capas de piedra arcillosa hasta un punto en el que todos ellos, con excepción de una única especie, desaparecían de modo repentino. Este punto de desaparición de los microorganismos marcaba el límite entre el cretáceo y el terciario, la era de la extinción masiva.
Se comprobó que ninguna inversión del campo magnético coincidía con las devastaciones ocurridas en el período limítrofe entre el cretáceo y el terciario. Pero donde los organismos desaparecían terminaba también la piedra caliza que era cubierta por una capa de tierra arcillosa de color gris rojizo de uno o dos centímetros de grosor. Esa tierra arcillosa estaba casi completamente desprovista de fósiles. Por encima de ella se prolongaba, de nuevo, la piedra caliza, llena de fósiles. Álvarez se sintió intrigado por la historia que podrían contar aquellas arcillas sobre la extinción de finales del cretáceo. Álvarez, catedrático asociado de Geología en la Universidad de California, en Berkeley, se llevó consigo algunas muestras de aquella arcilla cuando emprendió el regreso a su casa, en 1977, y describió a su padre lo que había encontrado. Su padre, Luis W. Álvarez era un físico que trabajaba en los Laboratorios Lawrence, en Berkeley, y entre cuyos múltiples honores se incluye un Premio Nobel. Los dos científicos, padre e hijo, se preguntaron asombrados cuántos años podían estar representados por aquella capa arcillosa que separaba dos formaciones de piedra caliza en relación con la vida en la Tierra. Si se conseguía ese conocimiento se podría llegar también a un entendimiento y comprensión de las extinciones del cretáceo-terciario. Aunque Luis Álvarez no era un geólogo se lanzó de lleno en el problema de la arcilla de Gubbio. Ése era el tipo de investigación que le iba bien. Se describía a sí mismo como un «cazador de choque», un físico subatómico que buscaba «resonancias». Cazando choques fue como detectó e identificó los rayos cósmicos, lo que constituía su interés principal. La arcilla de Gubbio enfrentó a Álvarez con un nuevo desafío. Lo afrontó desarrollando nuevos métodos de investigación científica. Concibió un método para medir la rapidez con que los sedimentos recogidos en el fondo del mar pasan a formar las capas de arcilla. Le recordó a su hijo Walter que dado que el polvo cósmico y el material micrometeorítico caen en el planeta a un ritmo más o menos constante, éste debe dejar una marca química con la cual se podría calcular la duración del período de tiempo en los límites del cretáceo. El material químico extraterrestre es químicamente diferente del que se encuentra en la superficie de la Tierra. El escaso elemento iridio, por ejemplo, es varios miles de veces superior en los meteoritos que en la corteza terrestre. Como el iridio tiene una afinidad por el hierro, como ocurre con otros miembros del grupo de elementos del platino, la mayor parte del iridio original debió de asentarse en el núcleo del planeta conjuntamente con el hierro fundido allí. Así Álvarez decidió medir la cantidad de iridio contenido en la arcilla. El estudio geoquímico de la arcilla de Gubbio, en el que el equipo de Álvarez estaba buscando iridio o cualquier otra materia extraterrestre reveladora, demostró cantidades tan escasas en la muestra que no podían suponer más de una parte por mil millones. Pero cuando Álvarez vio su primera «muestra» de iridio se dio cuenta de que el experimento para determinar el período de tiempo límite había fracasado, pues encontró en la arcilla mayor cantidad de iridio del que podría esperarse en base a la caída de polvo cósmico o del normal proceso de la Tierra.

En 1978, Walter Álvarez regresó a Gubbio para hacerse con una colección más completa de muestras de rocas buscada a 350 metros por debajo de la línea del límite del período. La concentración de iridio que había en ellas se vio que era notablemente constante mientras se iba subiendo en dirección a la franja límite, pero una vez allí aumentaba fuertemente. La capa de arcilla era treinta veces más rica en iridio que los sedimentos que se hallaban directamente por debajo o por encima de ella. Cuando los dos Álvarez, padre e hijo, vieron los resultados comenzaron a interrogarse sobre cuál podría ser su significado. No podían imaginarse que la existencia de una cantidad anómala de iridio fuera una coincidencia, precisamente allí, en la capa correspondiente al límite entre el cretáceo y el terciario, precisamente en el tiempo de la extinción masiva. No podían pensar en ningún proceso de depositación normal como factible explicación de lo que estaban viendo. Estaban seguros de que el iridio en la arcilla de Gubbio tenía que ser un jalón de significativa importancia en la historia de la Tierra. Walter Álvarez hizo su primer comunicado sobre sus descubrimientos en Gubbio en 1979, en una reunión de la Unión Geofísica Americana, en Washington. Dijo que las pruebas del iridio apoyaban las teorías en favor de que había sido un acontecimiento extraterrestre el que desató en la Tierra los cambios que llevaron a aquella extinción masiva. Al principio él y su padre creyeron que el material llegado a la Tierra procedía de una supernova. Cualquiera que fuera la fuente, si se trataba de una fuente extraterrestre, la intensidad del iridio parecía ser a nivel mundial como en realidad ocurría. El equipo de los Álvarez examinó muestras conseguidas en Dinamarca y otras de lugares tan alejados como Nueva Zelanda y pudo determinar que las concentraciones de iridio, inusualmente elevadas, estaban presentes siempre en la misma capa geológica con lo que no cabía duda de que se trataba de un fenómeno global. Finalmente los Álvarez llegaron a la conclusión de que el impacto de un meteorito, de un asteroide grande de al menos diez kilómetros de diámetro ofrecía un «escenario que explicaba la mayor parte, o toda, la evidencia biológica y física» asociada con la capa limítrofe y la extinción masiva. El asteroide, abriéndose paso por la atmósfera a 100.000 kilómetros por hora, pudo haber causado un cráter de 200 kilómetros de anchura y enviar a la estratosfera una nube de polvo que se extendería rápidamente hasta envolver por completo a la Tierra, sumiéndola en la oscuridad. Un desastre de mucha mayor magnitud tendría que haberse producido como consecuencia del impacto de un asteroide, tal y como lo veía el grupo Álvarez. No cabía duda de que el impacto tendría que haber provocado violentos terremotos y monumentales inundaciones en las costas. Tenía que haber lanzado al espacio un volumen de escombros sesenta veces mayor que la masa del propio asteroide. El polvo procedente de este fenómeno podría haber cambiado el día en noche durante varios años. Sin luz solar cesaría la fotosíntesis, lo que causaría la muerte de las plantas y dislocaría la cadena alimentaria desde su propia raíz. Hubiera sido un desastre sin paliativos. La extinción habría sido la suerte de muchas especies que vivían en la Tierra en el momento de producirse la colisión del asteroide.
Pero había muchos escépticos y algunos de ellos querían saber cómo era que el impacto del asteroide no había dejado un cráter conocido. Walter Álvarez comentó que nada le gustaría más que encontrar un cráter que pudiera ser el de su asteroide. Durante la década que siguió a la publicación del estudio, la hipótesis de la extinción por el choque de un asteroide continuó siendo tema de debate entre geólogos y paleontólogos. Uno de las mayores objeciones a esta hipótesis era que no se conocía un cráter cuyas dimensiones correspondieran al tamaño calculado, que debería tener entre 150 y 200 km de diámetro. Si bien no sería imposible que la Tierra hubiera cambiado desde entonces escondiendo una tal deformación, en 1990 se ubicaron indicios en Haití de un tsunami de grandes proporciones que arrastró residuos de iridio. Buscando estudios geológicos realizados desde los años 1960 en adelante se pudo ubicar un cráter en Chicxulub, en la península de Yucatán, con un diámetro de unos 170 km. Para algunos científicos, un problema de esta teoría es que la lectura de los registros fósiles sugiere que la extinción masiva de hace 65 millones de años duró cerca de diez millones de años, lo que no cuadra bien con que su causa fuera el impacto. Cuando el mesozoico se aproximaba a su fin, unos pocos dinosaurios de pequeño tamaño habían alcanzado una relación cerebro-peso corporal, o cociente de encefalización, que se podría comparar favorablemente con el de los mamíferos primitivos. Uno de esos dinosaurios era el Stenonychosaurus inequalus, una especie desenterrada por él en 1968, en unas capas geológicas del cretáceo, cerca del Parque Provincial de los Dinosaurios, en Alberta. Partiendo del cráneo y de algunos otros huesos recogidos, determinó que se había tratado de un carnívoro bípedo que debía de pesar unos cuarenta kilos con un metro de altura y que, con la cola extendida, podía medir desde su extremo a la punta del hocico unos dos metros. El tamaño de la cavidad craneal indicaba que elStenonychosaurus tenía un cociente de encefalización de 0,3, probablemente suficiente para hacer de él el más inteligente de los dinosaurios, con inteligencia varias veces superior a los demás, y posiblemente tan inteligente como los más avanzados de los pequeños mamíferos que fueron sus contemporáneos. En los seres humanos el cociente de encefalización es de 7,5. También había otros rasgos que hacían de este diminuto dinosaurio un ser excepcional. Al parecer tenía el pulgar libre, un dedo que podía oponerse a las otras dos garras, lo que sin lugar a dudas le daba una especial destreza y capacidad de manipulación. Es posible, también, que tuviera visión estereoscópica. «Parecía tener -dijo Russell- todos los componentes del éxito que posteriormente hemos visto en el desarrollo de los simios».
Con esta idea en mente, Russell se lanzó a formarse un concepto sobre el tipo de animal que hubiera llegado a ser el Stenonychosaurus en nuestros días si él y los demás dinosaurios no se hubieran extinguido hace sesenta y cinco millones de años. Dio forma a sus ideas en un modelo tridimensional de fibra de vidrio, creado con la ayuda de Ron Sequin, un taxidermista, y que en la actualidad se encuentra en el Museo de Ciencias Naturales en Otawa. Russell llama a este modelo un dinosauroide, una criatura inteligente que podría haber llegado a ser la culminación posible, en el siglo XX, de una evolución ininterrumpida de los dinosaurios, si éstos hubieran sobrevivido. El modelo de dinosauroide, según Russell, es el producto de un trabajo de suposiciones y una extrapolación «conservadora» de determinadas tendencias evolutivas. Tomando en consideración la tendencia hacia un mayor cerebro en los dinosaurios, que se había manifestado ya antes de su extinción, el dinosauroide hubiera evolucionado ahora hasta un cociente de encefalización del 7,1, que está dentro del límite de los humanos. Para dejar sitio al mayor cerebro, el cráneo en el modelo fue construido algo mayor que el delStenonychosaurus y el rostro relativamente más pequeño, como se vio ocurría en la evolución de los simios antropoides. Los ojos eran grandes óvalos, como es característico en los dinosaurios. La postura era erguida. El cuello más corto de lo que era corriente en los dinosaurios, porque de seguir con el cuello largo les hubiera resultado muy difícil soportar el peso de un cráneo grande al final de un cuello horizontal. La cola, al hacerse innecesaria como contrabalance para el equilibrio, habría desaparecido y por esa razón el dinosauroide no la tenía. Su cráneo es calvo y sus manos tienen tres dedos, ambas cosas características establecidas de los dinosaurios. Dado que Russell asume, además, que el animal habría evolucionado y dejado de poner huevos para traer al mundo a sus hijos vivos, el dinosauroide tenía ombligo. Russell creía que las crías serían alimentadas con alimentos previamente masticados o regurgitados, como lo son las crías de las aves, y por esa razón los dinosauroides no tenían pezones. La falta de órganos sexuales externos del dinosauroide es una característica típica de los reptiles. Cualquiera que fuese la fisiología y la conducta social de los dinosaurios en su época, concluía Russell, el dinosauroide habría evolucionado ya hasta convertirse en una criatura fuerte y vigorosa, de sangre caliente, que probablemente viviría formando comunidad con otros compañeros de caza. Estas criaturas podrían tener voces semejantes al piar de las aves y, posiblemente, algún tipo de lenguaje, aunque no hay modo de saber a qué nivel de encefalización el lenguaje se hace posible o inevitable. En el modelo de Russell, que como él dice es conservador, el dinosauroide aún no habría alcanzado niveles de inteligencia humana, pero el científico no excluye la posibilidad de que los descendientes del Stenonychosaurus, o cualquier otro de los dinosaurios mejor dotados cerebralmente, pudieran haber evolucionado hasta convertirse en especies más inteligentes, ingeniosas y sofisticadas que el Homo sapiens.
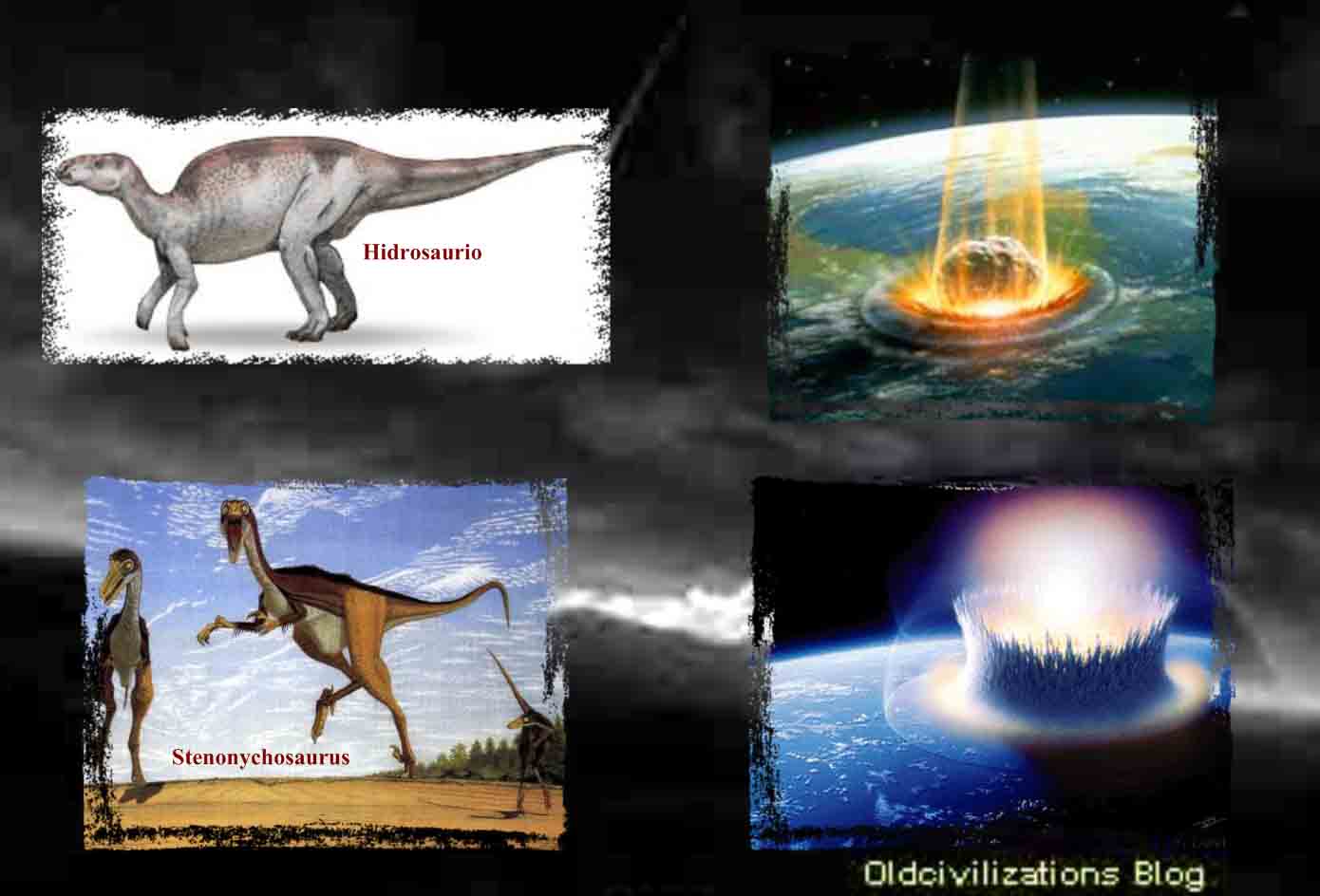
Pero se supone que el Stenonychosaurus nunca tuvo la posibilidad de llevar a cabo su hipotético potencial. Parece que ni un solo dinosaurio sobrevivió a las extinciones masivas de finales del cretáceo, ya que no se ha encontrado fósil alguno de esas criaturas en capas geológicas más recientes que las del cretáceo. Los informes ocasionales de que animales con aspecto de dinosaurios han sido vistos en África, nunca fueron confirmados. Aunque pudieran ser encontrados algunos animales monstruosos, con un curioso aspecto de dinosaurios, es altamente improbable que los verdaderos dinosaurios hubieran sobrevivido los últimos sesenta y cinco millones de años sin dejar ningún rastro fósil que hubiese sido descubierto hace ya muchos años. Todo ello si excluimos la remota posibilidad de que huyesen (o los llevasen) fuera de la Tierra. En la actualidad los dinosaurios viven sólo en las mentes de la especie que, sin duda, debe su existencia a la desaparición del dinosaurio. Si éste no se hubiera extinguido, si su género hubiese continuado dominando la tierra seca, es muy probable que los mamíferos se limitaran a ser pequeñas criaturas de aspecto de roedores, saliendo de noche y escondiéndose durante el día por temor a los poderosos reptiles. O, en el caso de que los dinosaurios hubieran vivido después del cretáceo para haberse extinguido en época más reciente, la Era de los Mamíferos posiblemente estaría aún en su infancia, sin que existieran todavía ni humanos ni protohumanos. Tenemos, pues, toda la razón al sentimos fascinados por los dinosaurios y, en especial, por su extinción. Las extinciones masivas, como los científicos están comenzando a reconocer, fueron acontecimientos cruciales en el curso de la vida en la Tierra. Tras la última extinción del cretáceo, debió de existir una enorme desolación por todas partes, silencio y calma donde antes hubo una vida fuerte y vigorosa. Pero, además, una oportunidad para todos aquellos supervivientes que tuvieran la suficiente capacidad de adaptación para aprovecharla. Las aves disponían del cielo para ellas solas, tras la extinción del pterosaurio. Los peces, los moluscos y otras criaturas del mar ya no tenían que vérselas con los ammonites ni con losmosasaurios. Los pequeños mamíferos salieron de su baja existencia; ya no quedaban saurios gigantescos, con la excepción de los cocodrilos, y éstos eran incapaces de mantener la hegemonía que durante tanto tiempo fue patrimonio de sus parientes arcosaurianos. Luis Álvarez se dio cuenta, rápidamente, del paralelismo existente entre la hipotética catástrofe que condenó a los dinosaurios y una guerra nuclear. En 1982 comparó de modo explícito el impacto de su supuesto asteroide con el potencial destructivo de las armas nucleares, una comparación que nos hace dudar de las esperanzas de que se pueda sobrevivir a una guerra atómica. Eso llevó a meditar a otros científicos. Si el impacto de un asteroide puede producir nubes de polvo globales y hacer bajar las temperaturas, ¿cuál sería el efecto de una guerra nuclear sobre el clima de nuestro planeta?
Fuentes:
John N. Wilford – El enigma de los Dinosaurios
Richard Leakey y Roger Lewin – La sexta extincion: el futuro de la vida y de la humanidad









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}